Регуляция мочеиспускания и дефикации

Высшие центры контроля мочеиспускания: цикл мочеиспускания, его нарушения
Нижние мочевыводящие пути разделены на два отдела-вместилище мочи (мочевой пузырь), а также собственно выводящие пути-уретру и наружный сфинктер уретры, действующих скоординировано для хранения и выведения мочи. На уровне спинного мозга парасимпатические нейроны крестцовых сегментов вызывают сокращение мочевого пузыря (детрузора). Симпатические нейроны поясничных сегментов угнетают парасимпатические влияния и позволяют мочевому пузырю наполниться.
Двигательные нейроны крестцовых сегментов, иннервирующие наружный сфинктер уретры, вносят вклад в функционирование обеих систем: при их возбуждении облегчается накопление мочи, при их торможении-облегчается ее выведение. Взаимодействие между этими системами происходит на различных уровнях ЦНС.
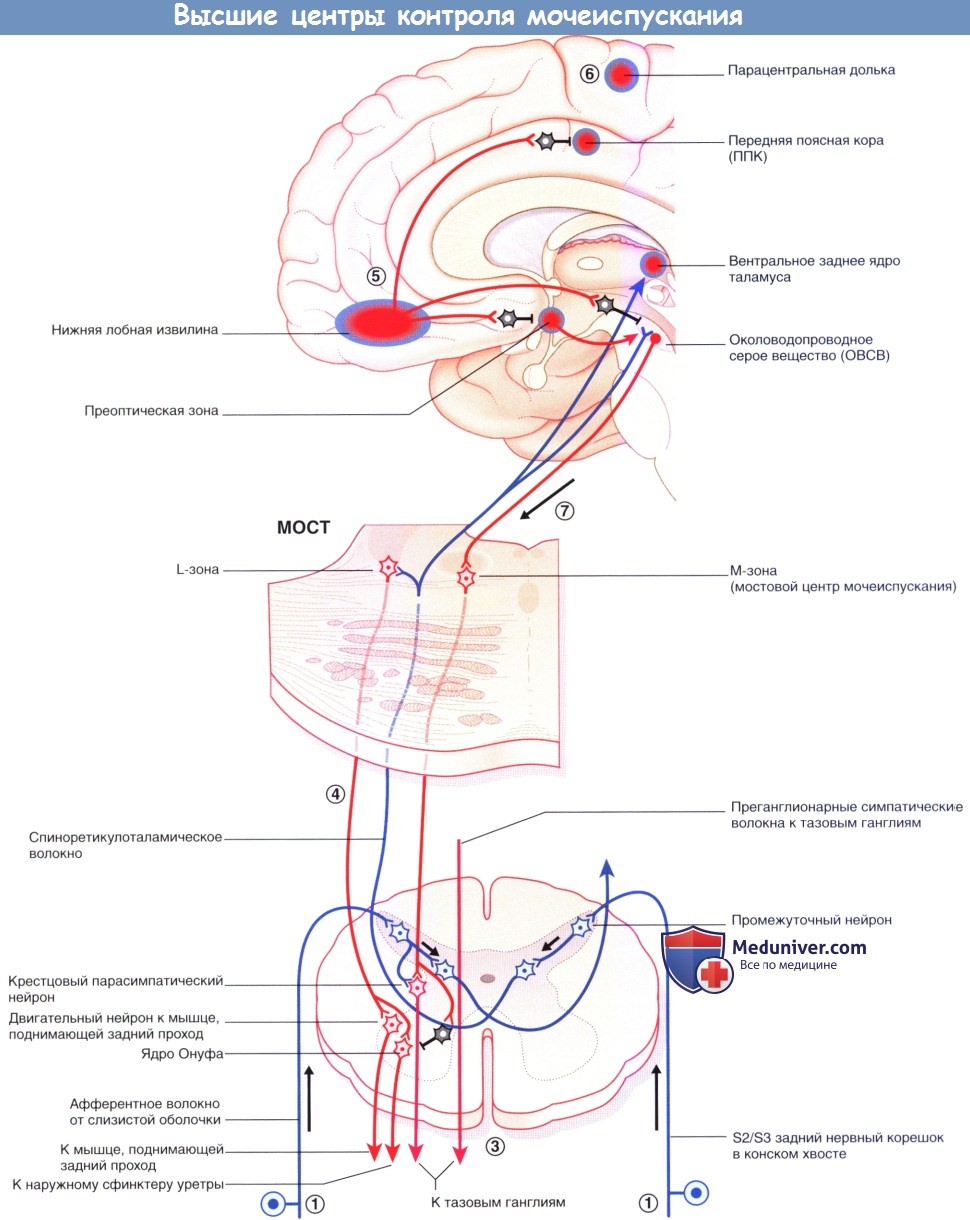
Мостовой центр контроля мочеиспускания (ядро Баррингтона) находится в парамедианной мостовой ретикулярной формации с каждой стороны, соединяясь по срединной линии. Отсюда отходят волокна крупноклеточных нейронов к отвечающим за мочеиспускание парасимпатическим нейронам в сегментах S2-S4 спинного мозга. Возбуждение центра контроля мочеиспускания приводит к акту мочеиспускания посредством повышения внутрипузырного давления (в результате сокращения гладких мышц стенки мочевого пузыря) и одновременного расслабления поперечно-полосатых мышц наружного уретрального сфинктера (вследствие возбуждения ГАМК-ергических вставочных нейронов ядра Онуфа, образующих синапсы с крестцовыми сегментами спинного мозга).
Аксоны этих двигательных нейронов проходят в составе половых нервов, иннервирующих наружный уретральный сфинктер. (В более латеральном отделе ретикулярной формации моста расположена L-зона, имеющаяся у млекопитающих, от которой волокна направляются к ядру Онуфа и вызывают сокращение наружного сфинктера уретры. Мостовой центр контроля мочеиспускания обозначают как М-зону, однако его взаимодействие с L-зоной вызывает сомнения.)
На более высоких уровнях к клеткам латерального отдела правого ОВСВ подходят восходящие волокна от задних рогов крестцовых сегментов, а от них-возбуждающие волокна к островку, который генерирует чувство наполненности мочевого пузыря и передает возбуждение на медиальные отделы лобной коры. Возбуждение латерального отдела ОВСВ осуществляет также правый гипоталамус. Часть спиноретикулярных волокон от крестцовых сегментов возбуждает L-зону. Другие волокна переключаются через таламус на клетки правой передней поясной коры (ППК), возбуждающиеся в ситуациях, требующих концентрации внимания.
Такое доминирование правых отделов может быть связано с эмоциональными аспектами мочеиспускания. При функциональном исследовании мозга было установлено, что дальнейшее заполнение жидкостью уже полного мочевого пузыря приводит к активации островка и дорсальной передней поясной коры/дополнительной моторной области. Когда мочевой пузырь не наполнен, заполнение жидкостью приводит к активации среднего мозга и парагиппокампальной области, осуществляющих бессознательный контроль афферентных сигналов от мочевого пузыря.
а) Цикл мочеиспускания:
1. Когда мочевой пузырь заполнен наполовину, восходящие волокна от рецепторов растяжения в детрузоре и слизистой оболочке в мочепузырном треугольнике передают эту информацию по спиноретикулярным волокнам через таламус к мосту, среднему мозгу и мозжечку.
2. От коры островка информация поступает в центр принятия решений в медиальной лобной коре, которая получает данные о степени наполнения мочевого пузыря.
3. Активация симпатической системы увеличивается постепенно, позволяя увеличить растяжимость мочевого пузыря (через β2-адренорецепторы). При взаимодействии с α2-адренорецепторами происходит торможение парасимпатических нейронов.
4. Спиноретикулярные волокна, образующие синапсы с L-зоной моста, возбуждают ядро Онуфа в крестцовых сегментах спинного мозга, повышая, таким образом, тонус наружного сфинктера уретры.
5. При полном заполнении возникает ощущение непреодолимого желания. При неподходящем времени или месте активируется отдел медиальной лобной извилины. Эта область тормозит ППК, уменьшая уровень ее активности путем возбуждения ее тормозных вставочных нейронов. Аналогичным образом через переключение в гипоталамусе и среднем мозге происходит торможение пре-оптической области и ОВСВ посредством возбуждения тормозных нейронов.
6. Последняя, но кратковременная мера -произвольное сокращение мышц тазовогодна. Команду на сокращение посылает лобная кора к промежностному отделу в медиальной области моторной коры парацентральной дольки.
7. Когда время и место позволяет, медиальная лобная извилина «освобождает троих своих пленников». Происходит расслабление тазового дна, гипоталамус активирует М-зону ОВСВ, одновременно с этим ингибируя L-зону путем возбуждения тормозных вставочных нейронов. Правостороннее доминирование центра контроля мочеиспускания согласуется с клиническими данными: у пациентов обоих полов, перенесших инсульт, недержание мочи чаще возникает при правосторонней локализации очага.
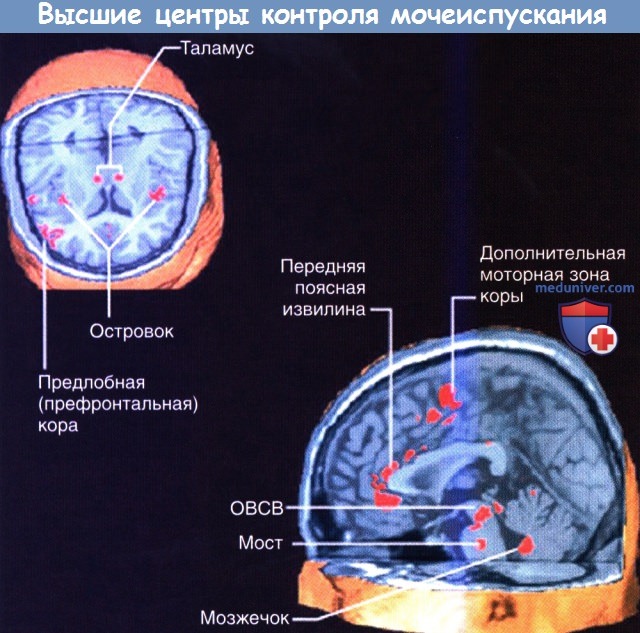 Показаны высокоактивные области при функциональной магнитно-резонансной томографии (фМРТ) в фазу наполнения мочевого пузыря в цикле мочеиспускания.
Показаны высокоактивные области при функциональной магнитно-резонансной томографии (фМРТ) в фазу наполнения мочевого пузыря в цикле мочеиспускания.
ОВСВ – околоводопроводное серое вещество. Мост – подразумевается мостовой центр контроля мочеиспускания.
б) Роль моноаминов. К двигательным и чувствительным ядрам спинного мозга, иннервирующим мочевой пузырь, подходит огромное количество серотонинергических нисходящих волокон от большого ядра шва (БЯШ) продолговатого мозга. Доказано, что растяжение мочевого пузыря приводит к стимуляции БЯШ (путем спиноретикулярной активации ОВСВ). Кратко излагая, контроль мочеиспускания на низшем уровне означает, что БЯШ создает тонус мочевого пузыря, направленный, прежде всего, на его наполнение.
Норадренергические нисходящие волокна из голубого пятна к передним рогам спинного мозга усиливают действие глутамата, локально вырабатываемого из клеток ядра Онуфа, увеличивая, тем самым, тонус наружного сфинктера в фазу наполнения.
в) Императивное недержание мочи. Императивное недержание мочи – неспособность взрослой женщины контролировать опорожнение мочевого пузыря до завершения фазы наполнения цикла мочеиспускания. Оно характеризуется неотложным позывом, быстро переходящим в неконтролируемое мочеиспускание, независимо от обстоятельств (отсюдатермин – «гиперактивный мочевой пузырь», или «гиперактивность детрузора»), Во многих случаях в анамнезе имеется указание на раздраженный мочевой пузырь в детском возрасте в форме частого дневного мочеиспускания и/или ночного энуреза (ночного недержания мочи).
При функциональном исследовании мозга (фМРТ) у здорового взрослого человека можно обнаружить повышение активности правой островковой области, которая, вероятно, отвечает за повышенную реактивность мочевого пузыря, и передней (отвечающей за эмоции) области поясной коры, обусловливающей возникновение неотложного позыва и «страх» неизбежного мочеиспукания.
G-белковые мускариновые рецепторы, активируемые постганглионарными волокнами тазовых ганглиев, в большом количестве присутствуют в детрузоре мочевого пузыря. Соответственно, препаратами выбора в данном случае служат блокаторы м-холинорецепторов. Однако появление антихолинергических побочных эффектов, таких как сухость во рту и запор, может потребовать отмены терапии.
В последние годы для лечения гиперактивности детрузора все чаще используют ботулотоксин. Известно, что он нарушает взаимодействие между холинергическими синаптическими везикулами и мышечными волокнами (как гладкими, так и поперечно-полосатыми), делая синаптическую передачу неэффективной. Гибкий цистоскоп проводят через уретру и выполняют многочисленные небольшие инъекции ботокса в стенку мочевого пузыря. Для получения долговременного результата процедуру повторяют 2 раза в год.
– Вернуться в оглавление раздела “Неврология.”
Редактор: Искандер Милевски. Дата публикации: 20.11.2018
Источник
^Нервная регуляция опорожнения мочевого пузыряМоча, постоянно вырабатывающаяся в почках, накапливается в мочевом пузыре, который периодически полностью опорожняется. Эта функция, играющая огромную роль в социальной жизни человека, опосредована как деятельностью гладких мышц мочевого пузыря, так и влияниями вегетативных и соматических нервов. Нервная регуляция функции мочевого пузыря заключается в чередова- ^ ЧАСТЬ IV. ПРОЦЕССЫ НЕРВНОЙ И ГУМОРАЛЬНОЙ РЕГУЛЯЦИИ
нии длительных периодов наполнения и коротких периодов опорожнения. Благодаря нервной активности опорожнение мочевого пузыря во время периода наполнения невозможно или затруднено. Скорость наполнения мочевого пузыря составляет примерно 50 мл в час. Вследствие пластичности гладких мышц пузыря (см. с. 86) давление в нем лишь незначительно повышается при увеличении объема (рис. 16.15). При накоплении приблизительно 150-250 мл мочи появляются первые короткие позывы к мочеиспусканию, обусловленные кратковременным повышением давления внутри пузыря. Период опорожнения обычно начинается, когда в мочевом пузыре накапливается порядка 250-500 мл мочи. Способность пузыря накапливать мочу называется удержанием мочи, а его опорожнение-актом мочеиспускания (рис. 16.15). Строение и иннервация мочевого пузыря (рис. 16.16). Мочевой пузырь представляет собой полый орган, образованный мышечными слоями; эти слои в совокупности называются изгоняющей мышцей (детрузором). Стенки пузыря состоят из сети длинных гладкомышечных волокон. В области основания пузыря находится треугольный участок, образованный тонкими гладкомышечными волокнами (пузырный треугольник). В углах основания этого треугольника расположены устья мочеточников. Мочеточники открываются в пузырь в косом направлении, поэтому при повышении внутрипузырного давления обратного заброса мочи в них не происходит. В области вершины треугольника отходит мочеиспускательный канал. Благодаря особому расположению мышц здесь образуется как бы функциональный сфинктер (внутренний сфинктер мочевого пузыря). При акте мочеиспускания внутренний сфинктер может расслабиться только в результате сокращения детрузора; вследствие сокращения этой мышцы мочеиспускательный канал под действием вплетающихся в него радиальных волокон укорачивается, что автоматически приводит к пассивному раскрытию внутреннего сфинктера. Мочеиспускательный канал замыкается, кроме того, наружным сфинктером, образованным поперечнополосатой мускулатурой промежности; у женщин этот сфинктер выражен слабо. Иннервация мочевого пузыря и сфинктеров схематически представлена на рис. 16.16. ^ тазовых нервов, отходящие от второго и четвертого крестцовых сегментов, возбуждают мускулатуру пузыря. Влияние этих нервов необходимо для нормальной регуляции акта мочеиспускания. Под действием симпатических нервов, отходящих от верхних поясничных сегментов, мышца, изгоняющая мочу, расслабляется, а мышцы пузырного треугольника сокращаются. Возможно, эти симпатические влияния способствуют удержанию мочи. Соматическая иннервация наружного сфинктера осуществляется двигательными волокнами срамного нерва, тела нейронов которых лежат в средних крестцовых сегментах. Информация о степени наполнения мочевого пузыря воспринимается рецепторами растяжения, расположенными в его стенке, и передается в центральную нервную систему по чувствительным волокнам тазовых нервов. Возможно, болевые и неболевые раздражения, возникающие при различных процессах в мочевом пузыре и мочеиспускательном канале, передаются в виде импульсов как в крестцовых, так и в поясничных висцеральных афферентах.
^
^ Из почечных лоханок моча поступает в мочевой пузырь благодаря перистальтическим сокращениям мочеточников. Чем более растянута стенка мочевого пузыря, тем сильнее возбуждаются расположенные в ней рецепторы растяжения. Возбуждение этих рецепторов приводит к активации парасимпатических нейронов, иннервирующих детрузор (рис. 16.17, рефлекторная дуга 1). В результате мочевой пузырь опорожняется. Как указано на рис. 16.17, дуга этого рефлекса замыкается на уровне передней области моста. Электрическое раздражение этой области приводит к опорожнению мочевого пузыря. С началом опорожнения пузыря процесс ускоряется лавинообразно до полного изгнания мочи. Этот протекающий по принципу положительной обратной связи (т.е. усиливающий сам себя) процесс, возможно, обусловлен следующими основными рефлексами: 1) сокращение мышцы, изгоняющей мочу, усиливает возбуждение афферентных волокон от рецепторов пузыря; 2) под влиянием струи мочи возбуждаются чувствительные волокна от рецепторов мочеиспускательного канала, что приводит к рефлекторной активации парасимпатических нервов мочевого пузыря; 3) происходит рефлекторное угнетение тормозных процессов в спинном мозге и вышележащих отделах. Все это сопровождается также рефлекторным расслаблением наружного сфинктера в результате торможения мотонейронов крестцовых сегментов. Как у животных, так и у людей перерыв спинного мозга выше крестцовых сегментов вначале приводит к полному подавлению рефлекторного опорожнения мочевого пузыря. Лишь по достижении хронической стадии заболевания (1-5 недель после травмы) вновь начинает действовать рефлекс опорожнения (автоматическое опорожнение). В этом случае рефлекторная дуга замыкается только в пределах спинного мозга (рис. 16.17, рефлекторная дуга 2). Вполне вероятно, что эта же дуга регулирует мочеиспускание у грудных детей; с возрастом же, по-видимому, этот рефлекс подавляется в результате тормозных влияний как от афферентных пузырных волокон, замыкающихся на уровне спинного мозга, так и от супраспинальных отделов. В течение первых дней или недель после возникновения параплегии или тетраплегии наблюдается вялость и атоничность мочевого пузыря. Если благодаря правильному уходу за больным не добавится инфекция мочевых путей, то начинается постепенный переход к автоматическому опорожнению мочевого пузыря. В этот период незначительное наполнение пузыря приводит к рефлекторному сокращению детрузора; мочеиспускание при этом учащено. В результате соответствующей тренировки такие больные могут управлять опорожнением пузыря. Они обучаются вызывать рефлекторные сокращения мышцы, изгоняющей мочу; для этого больной в определенное время, приуроченное к автоматическому ритму мочеиспускания, постукивает по нижнему отделу живота и надавливает на брюшную стенку, способствуя тем самым опорожнению пузыря (сегментарный рефлекс; см. с. 357). ^ Регуляция мочеиспускания и удержания мочи представляет собой в значительной степени автоматический рефлекторный процесс, однако на него оказывают влияние центры высших отделов ствола мозга, гипоталамус и большие полушария. Все эти влияния преимущественно ингибиторные, хотя некоторые из них могут быть и возбуждающими. О восходящих и нисходящих спинальных путях, соединяющих мочевой пузырь и мочеиспускательный канал с высшими отделами ЦНС, а также о расположении соответствующих нейронов в стволе мозга, гипоталамусе и коре больших полушарий известно мало. Деятельность высших отделов заключается, во-первых, в удержании мочи даже при сильном наполнении мочевого пузыря (это необходимо для того, чтобы мочеиспускание происходило только в надлежащих условиях) и, во-вторых, в произвольном мочеиспускании при наличии соответствующих позывов и условий [7, 11, 38]. Нарушения мочеиспускания весьма разнообразны и широко распространены. Непроизвольная задержка мочи может наступать при параличе или повреждении детрузора (например, при воспалительных заболеваниях или травматических повреждениях нервов), смешении мочеиспускательного канала (например, из-за опухоли предстательной 364 ^ железы) или при спазмах сфинктеров. Неспособность удерживать мочу называется недержанием. Она особенно часто встречается у рожавших женщин (например, в результате опущения матки из-за слабости мышц дна таза), а также при органических поражениях мозга (например, при рассеянном склерозе или атеросклерозе сосудов мозга у пожилых людей). Встречается также психогенное недержание мочи [16]. ^ Опорожнение кишечника (дефекация) и удержание каловых масс представляют собой важнейшие функции прямой кишки и ануса. Эти функции регулируются энтеральной нервной системой, парасимпатическими нервами из крестцовых сегментов и соматомоторными нервными влияниями. Внутренний анальный сфинктер имеет плотную симпатическую иннервацию, однако о роли симпатических нервов в функции низших отделов кишечника известно мало. Удержание кала. В области дистального конца прямой кишки имеются два сфинктера. ^ образованный гладкими мышцами, управляется непроизвольно. Наружный анальный сфинктер состоит из поперечнополосатых мышечных волокон, иннервируемых мотонейронами крестцового отдела (S2-S4), отростки которых идут в составе тазовых нервов. В покое оба этих сфинктера замкнуты. Тоническое сокращение наружного сфинктера поддерживается в результате спинальных рефлексов, афферентное звено которых включает импульсы от мышечных волокон сфинктера и окружающих тканей, и особенно кожи области анального отверстия. Когда в результате перистальтических сокращений нисходящей толстой кишки каловые массы поступают в прямую кишку, стенки ее растягиваются, что приводит к расслаблению внутреннего анального сфинктера и усилению сокращения наружного сфинктера. Расслабление внутреннего сфинктера связано главным образом с рефлексом, замыкающимся в пределах энтеральной нервной системы. Сокращение наружного сфинктера также возникает рефлекторно под влиянием сигналов, поступающих в крестцовые отделы спинного мозга по афферентным волокнам тазовых нервов (рис. 16.18). Эти процессы сопровождаются появлением позывов к дефекации – осознанных ощущений, связанных с возбуждением рецепторов стенок толстой и прямой кишок. Через несколько десятков секунд внутренний сфинктер вновь постепенно сокращается и наступает адаптация стенок прямой кишки к ее увеличенному объему. Эта адаптация обусловлена пластичностью ректальной мускулатуры. Напряжение в стенке прямой кишки снижается, и позыв к дефекации стихает. Благодаря всем этим процессам, находящимся под
контролем нервных влияний, здоровый человек может сдерживать позывы к дефекации до тех пор, пока объем содержимого прямой кишки не превысит 2 л. Важнейшую роль в удержании каловых масс играют супраспинальные центры, и особенно кора головного мозга: эти отделы оказывают возбуждающее влияние на мотонейроны наружного сфинктера и, возможно, ингибирующее влияние на парасимпатические спинальные рефлексы. Дефекация. Опорожнение прямой кишки в норме совершается произвольно. Под действием супраспинальных облегчающих влияний на спинальные парасимпатические рефлексы нисходящая, сигмовидная и прямая кишка (особенно их продольная мускулатура) сокращаются. Одновременно наступает расслабление обоих сфинктеров. Важным условием дефекации является повышение внутрибрюшного давления под действием, во-первых, усиленных сокращений мышц брюшной стенки и, во-вторых, опущения диафрагмы, наступающего в результате сокращения мышц грудной клетки, находящейся в положении вдоха, при замкнутой голосовой щели. Все эти механизмы приводят к тому, что тазовое дно опускается и каловые массы, содержащиеся в нисходящей, сигмовидной и прямой кишке, выбрасываются. Рефлексы дефекации полностью исчезают при разрушении крестцовых сегментов спинного мозга. После перерыва спинного мозга выше этих сегментов спинальные рефлексы дефекации сохраняются, однако произвольные движения, способствующие изгнанию каловых масс, не могут осуществляться. ^ Правда, они могут быть компенсированы (например, расширением наружного анального сфинктера руками), и благодаря этому больные параплегией способны регулярно опорожнять кишечник [7, 50]. | ||||||||
| Физиология человека. В 3-х томах. Т. Пер с англ./ под ред. Р. Шмидта и Г. Тевса. М.: Мир, 1996. 198 с Физиология человека: в 3-х томах / пер с англ под ред. Р. Шмидта и Г. Тевса. – М.: Мир. 1996.] | Руководство по терапевтической гомеопатии: Пер с англ. М.: Атлас, 1994. 205 с Аллен Х. К. Основания и показания к назначению и характеристики ведущих гомеопатичекских препаратов и нозодов со сравнением их патогенезов:… | |||||||
| Література … | Глоссарий Глоссарий взят из книги Брамм П., Брамм Д. Микропроцессор 80386 и его программирование: Пер с англ. М.: Мир, 1990,-448 с. Абсолютный адрес Глоссарий взят из книги Брамм П., Брамм Д. Микропроцессор 80386 и его программирование: Пер с англ. М.: Мир, 1990,-448 с | |||||||
| Пустыльник Е. И. Статистические методы анализа обработки наблюдений Базара М., Шетти К. Нелинейное программирование. Теория и алгоритмы: Пер с англ. М.: Мир, 1982. – 583 С | Список рекомендованої літератури Дункан Д. У. Основополагающие идеи в менеджменте. Уроки основоположников менеджмента и управленческой практики / Пер с англ. – М.:… | |||||||
| Список рекомендованої літератури Дункан Д. У. Основополагающие идеи в менеджменте. Уроки основоположников менеджмента и управленческой практики / Пер с англ. – М.:… | О.І. Рогач [та ін.]; pа ред. О.І. Рогача. К.: Либідь,2003. 784с. Isbn 966-06-0293-6 Мировые финансы[Текст]: пер с англ./ М. В. Энг, Ф. А. Лис, Л. Д. Мауер. М.: ДеКА,1998. 736с. Алф указ.: с. 722-734. Isbn 5-89645-004-4(рус.):… | |||||||
| Учебник для вузов системы мвд/ : в 2-х кн под ред проф. А. А. Пушкина (Олександр Анатолійович). Х.: Основа,1996 Цивільне право України : Підручник : у 2-х кн. /за ред. О. В. Дзери, Н. С. Кузнєцової-К.: Юрінком Інтер, 2001; 2002, 2004 | Руководство по коксованию том пер с немецкого Продукты каменноугольной смолы из книги: “Руководство по коксованию” том пер с немецкого под ред. О. Гросскинского с. 437 М.: Металлургия,… |
Источник