Мочевой пузырь рыбы функции

Основной функцией выделительной системы любого живого существа, в том числе и рыбы, является вывод из организма продуктов обмена веществ и сохранение водно-солевого баланса в крови и тканях. Разумеется, выделительная система рыб имеет более простое строение, чем, к примеру, человеческая. Выполнение функций происходит по определенной цепочке, для понимания которой следует изучить строение системы в целом и работу ее органов в отдельности.
Строение: какие органы образуют выделительную систему рыбы
За вывод из организма ненужных, а зачастую и ядовитых веществ, у этих представителей водной фауны, как и у человека, отвечают парные почки, представляющие собой сложную систему мелких проводных канальцев. Последние открываются в общий выводной проток. Мочевой пузырь у большинства рыб выходит наружу отдельным отверстием.
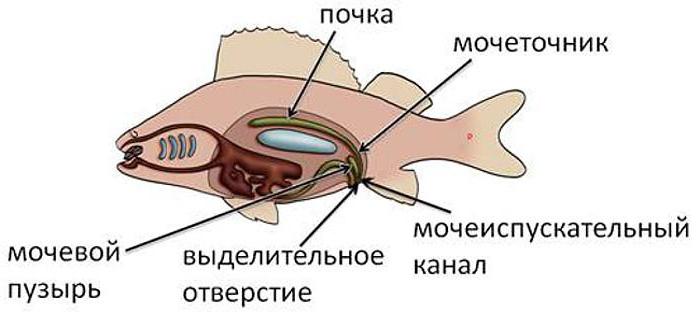
Образующиеся в почках продукты обмена через протоки преимущественно поступают в мочевой пузырь.
Почки мальков
Понимая, какие органы образуют выделительную систему рыбы, можно сделать вывод, что ключевая роль в ее функционировании принадлежит почкам.
В эволюционной цепочке рыбы занимают далеко не первое место. Биологи относят их к классу низших позвоночных. По сложности строения органов водоплавающие уступают и земноводным, и рептилиям. У высших позвоночных, в том числе человека, почки тазовые. У рыб они туловищные.
Степень сложности строения почек у любых живых существ определяется:
- количеством канальцев;
- наличием и строением мерцательных воронок.
У некоторых представителей фауны почки закладываются в верхней части и состоят из 6-7 канальцев. Мерцательная воронка, выполняющая роль фильтра, у таких организмов одним концом открывается в мочеточник, другим — в полость тела. Именно таким строением характеризуются почки мальков и некоторых взрослых рыб. К таковым относятся бельдюга, атерина, бычки и прочие. У других разновидностей рыб примитивная почка постепенно преобразуется в лимфоидный кровотворящий орган.
Почки взрослых рыб
У мальков в большинстве случаев почка расположена в верхней части туловища. У взрослых рыб этот парный орган заполняет пространство между плавательным пузырем и позвоночником. Как уже упоминалось, почки этих представителей водной стихии относятся к классу туловищных и выглядят в виде лентовидных тяжей темно-бордового цвета.
Основным функциональным элементом почки взрослой рыбы является нефрон. Последний в свою очередь состоит из:
- выделительных канальцев;
- мальпигиева тельца.
Мальпигиево тельце у рыб образуется капиллярным клубочком и капсулами Шумлянского — Боумена, представляющими микроскопические чашечки с двойными стенками. Мочевые канальцы, отходящие от них, открываются в собирательные. Последние, в свою очередь, сливаются в более крупные и выпадают в мочеточники.
Мерцательные воронки в почках большинства рыб отсутствуют, кроме некоторых видов. Такие функциональные элементы, к примеру, имеются у осетровых и некоторых хрящевых.
Примеры строения
Почки — довольно сложные по строению органы выделительной системы рыб. Принято выделять три основных отдела:
- передний (головная почка);
- средний;
- задний.
Отделы почек разных видов рыб могут иметь неодинаковую форму. Рассмотреть строение этого органа конкретно для каждого класса в одной небольшой статье, к сожалению, довольно сложно. Поэтому в качестве примера разберемся, как выглядит почка сазана, щуки и окуня. У карповых правая и левая почки расположены по отдельности. Ниже они соединяются в непарную ленту. Хорошо развитый средний отдел сильно расширен и в виде ленты охватывает плавательный пузырь.
У окуня и щуки почки имеют немного другое строение: средние отделы расположены порознь, а передние и задние соединены.
Мочевой пузырь
Строение выделительной системы рыб достаточно сложное. Мочевой пузырь имеется у большинства разновидностей этих представителей водной фауны.
Основных классов рыб в природе, как известно, встречается всего два:
- хрящевые;
- костные.
Различие между ними, в первую очередь, заключается в строении скелета. В первом случае он состоит из хрящей, во втором — соответственно, из костей. Класс хрящевых рыб представлен в природе примерно 730 видами. Костных представителей водной фауны гораздо больше: порядка 20 тыс. разновидностей.
Выделительная система рыб (костных и хрящевых) имеет неодинаковое строение. У первых имеется мочевой пузырь, а у вторых его нет. Конечно же, отсутствие этого органа у хрящевых рыб вовсе не означает того, что их ВС несовершенна. Свои функции она выполняет просто отлично.
Выделительная система хрящевых рыб включает в себя органы, строение которых максимально препятствует бесконтрольному поступлению мочи в окружающую среду. «Жидких отходов» в воду такие представители фауны выделяют обычно очень мало.
Ректальная железа рыб
Как уже упоминалось, выделительная система рыб отвечает не только за вывод продуктов обмена веществ, но и за сбережение в организме нормального уровня водно-солевого баланса. У рыб эту функцию выполняет ректальная железа, представляющая пальцеобразный вырост, который отходит от спинной части прямой кишки. Железистые клетки ректальной железы выделяют особый секрет, содержащий в себе большое количество NaCl. В первую очередь этот орган удаляет из организма избыточную соль, поступающую с пищей или морской водой.
Помимо сохранения солевого баланса, ректальная железа рыб выполняет еще одну очень важную функцию. В период размножения выделяемая слизь тянется вслед за рыбой, привлекая характерным запахом особей противоположного пола.
Солевой баланс
Осмотическое давление у всех подобных представителей фауны (как морских, так и пресноводных) значительно отличается от показателей окружающей среды. Единственным исключением из этого правила являются миксины. Концентрация солей в их организме такая же, как и в морской воде.
У хрящевидных рыб, относящихся к группе изоосмотических, давление такое же, как и у миксин и совпадает с давлением воды. Но концентрация солей при этом на порядок ниже, чем во внешней среде. Баланс давления в рыбьем организме обеспечивается высоким содержанием мочевины в крови. Концентрирование и удаление хлоридов-ионов и ионов натрия из организма производится ректальной железой.
Выделительная система костных рыб хорошо приспособлена для регулировки солевого баланса. Давление у таких представителей фауны регулируется немного по-другому. К классу изоосмотических такие рыбы не относятся. Поэтому в процессе эволюции они выработали особые механизмы, контролирующие и регулирующие содержание солей в крови.
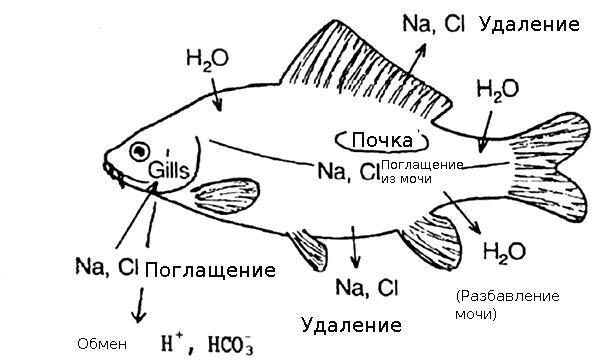
Так, морские костные рыбы постоянно теряющие воду под воздействием осмотического давления, для компенсации потерь вынуждены очень часто пить. Морская вода в их организме постоянно отфильтровывается от солей. Последние выводятся из организма двумя путями:
- катионы кальция с хлорид-ионами выбрасываются через жаберные мембраны;
- катионы магния с сульфатными анионами выводятся почками.
У костных пресноводных рыб, в отличие от морских, концентрация солей в организме ниже, чем во внешней среде. Выравнивают представители фауны давление путем захвата ионов из воды через жаберные мембраны. К тому же в организме таких холоднокровных вырабатывается большое количество мочевины.
Состав мочи
Как мы выяснили, по строению выделительная система рыб (хрящевых и костных) несколько различается. Неодинакова по составу и моча этих представителей фауны. Основным компонентом жидких выделений костных рыб является аммиак – вещество, токсичное даже в минимальных концентрациях. У хрящевых — это мочевина.

Доставляются продукты обмена веществ в почки рыб, по сути являющиеся фильтраторами, с током крови. Последняя предварительно подается в сосудистые клубочки. Именно в них и происходит процесс фильтрации, в результате которого формируется первичная моча. Сосуды, выводимые из клубочков, опутывают выделительные канальцы. Соединяясь вместе, они формируют задние кардинальные вены.
В средней части канальцев (в почках) происходит образование вторичной (окончательной) мочи. Здесь, помимо всего прочего, происходит всасывание веществ, необходимых организму. Это может быть, к примеру, глюкоза, вода, аминокислоты.
Пронефрический канал
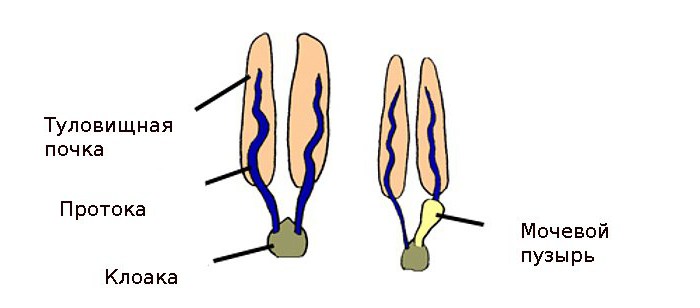
Выделительная система рыб представлена пронефрическим каналом – основным выходным протоком главной почки. У хрящевых рыб он состоит из двух частей: вольфов и мюллеров каналы. Последний имеется только у самок. У самцов он атрофирован.
У мальков вольфов канал предназначен для выполнения функций семяпровода. У самца хрящевой разновидности по мере взросления образуется отдельный мочеточник, который открывается в мочеполовой синус. Последний, в свою очередь, соединен с клоакой. У взрослых особей в семяпровод трансформируется вольфов канал.
Особенности выделительной системы рыб костных видов — это, прежде всего, отсутствие клоаки и разъединенность выводящей и половой систем. Вольфовы каналы у таких представителей фауны объединены в непарный поток. Последний при этом располагается по стенке брюшной полости рыбы сзади, образуя на пути мочевой пузырь.
Источник
Мочеполовая система рыб представлена органами мочевыделительной и репродуктивной систем. Органы этих двух систем тесно связаны между собой, поэтому их часто описывают вместе, хотя выделительные и половые каналы у рыб разделены.
К данным органам относятся: почки, половые железы (гонады), выводные протоки и наружные мочевыводящие и половые органы.
Почки у рыб обычно парные, состоящие из тёмно–красных тел лентовидной формы. Почки расположены почти вдоль всей полости тела и плотно прижаты к позвоночнику. Внутри почки находится мочевой канал, пронизанный капиллярами. Почка отфильтровывает из крови продукты распада, затем они попадают в этот канал, а кровь выходит из почек уже очищенная.
Кроме этого почки поддерживают и физико-химическую устойчивость организма: осмотическое давление и кислотно-щелочное, ионное равновесие.
Почки морских рыб задерживают воду; пресноводных — наоборот, откачивают ее из организма. Поэтому объем мочи у пресноводных рыб в 10 раз больше чем у морских. Колюшка сооружает место для нереста из растений, укрепляя их с помощью почечного секрета, быстро затвердевающего в воде.
Продукты распада из мочевого канала попадают в мочевой пузырь и выводятся наружу через мочеточник. Однако у некоторых рыб мочеточник выходит в анальное отверстие, из которого моча уже и выводится наружу. Кроме того, встречаются и виды рыб, у которых мочеточник выполняет одновременно и функцию семяпровода при размножении.
Оплодотворение у рыб может быть как внутренним (при помощи копулятивных органов), так и внешним: выметывание икры.
Гонады (семенники и яичники) рыб — это обычно парные лентовидные или мешковидные образования, подвешенные на складках брюшины: брыжейке, в полости тела рыбы.
По форме, гонады могут быть разными, например, у некоторых видов встречаются полностью слитые в одну железу (например, окунь), ассиметричные парные гонады. Встречаются и виды с одиночными (непарными) гонадами (карась серебряный).
Половой орган самок рыб — яичники. В них созревает и скапливается икра.
Яичники рыбы сливаются с яйцеводом (мюллеровым каналом), который выводит икру наружу. Некоторые виды (корюшковые, лососёвые, угрёвые) имеют незамкнутые яичники, и созревшие икринки попадают в полость тела, после чего через специальные каналы выводятся из организма. У живородящих рыб молодь развивается в яичниках.
Половой орган самцов рыб — семенник. В нем созревает и накапливается сперма. Наружу она выходит через семяпровод (вольфов канал) и половое отверстие (у самцов щуки, лососей), а у некоторых видов и через мочеполовое отверстие (у самцов большинства костистых рыб).
Строение и функции половых протоков, как и гонад, у разных видов рыб может быть разным.
По мере роста и полового созревания рыбы меняются размеры и внешний вид гонад.
Существует шкала зрелости гонад, пользуясь которой по внешним признакам (размерам и внешнему виду), устанавливают стадию созревания половых продуктов рыбы. Это очень важно при промышленном, декоративном и научном разведении рыбы.
Существуют и рыбы с принципиально иным строением мочеполовой системы — это например рыбы-гермафродиты (морской окунь).

Источник
Анисимова И.М., Лавровский В.В.
“Ихтиология”
Из-во Высшая школа. 1983 г.
ГЛАВА I
СТРОЕНИЕ И НЕКОТОРЫЕ ФИЗИОЛОГИЧЕСКИЕ ОСОБЕННОСТИ РЫБ
ВЫДЕЛИТЕЛЬНАЯ СИСТЕМА И ОСМОРЕГУЛЯЦИЯ
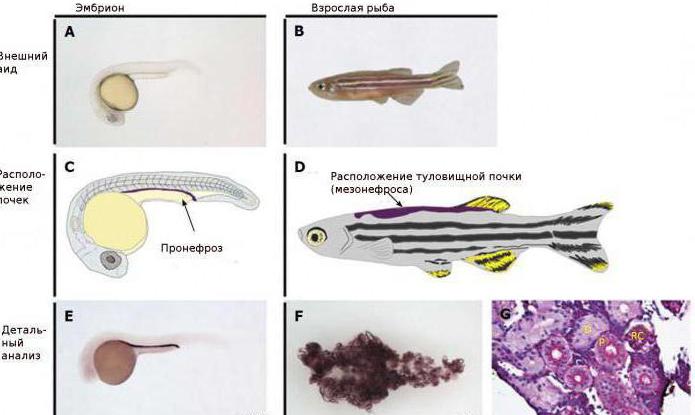
В отличие от высших позвоночных, имеющих компактную тазовую почку (метанефрос), рыбы обладают более примитивной туловищной почкой (мезонефрос), а их зародыши – предпочкой (пронефрос). У некоторых видов (бычок, атерина, бельдюга, кефаль) предпочка в том или ином виде выполняет выделительную функцию и у взрослых особей; у большинства же взрослых рыб функционирующей почкой становится мезонефрос.
Почки – парные, вытянутые вдоль полости тела темно-красные образования, плотно прилегающие к позвоночнику, над плавательным пузырем (рис. 22). В почке выделяют передний отдел (головная почка), средний и задний.
Артериальная кровь поступает в почки по почечным артериям, венозная по воротным венам почек.
Рис. 22. Почка форели (по Строганову, 1962):
1 – верхняя полая вена, 2 – выносящие почечные вены, 3 – мочеточник, 4 – мочевой пузырь
Морфофизиологическим элементом почки является извитой почечный мочевой каналец, один конец которого расширяется в мальпигиево тельце, а другой отходит к мочеточнику. Железистые клетки стенок секретируют продукты азотистого распада (мочевину) , которые попадают в просвет канальцев. Здесь же, в стенках канальцев, происходит обратное всасывание воды, сахаров, витаминов из фильтрата мальпигиевых телец.
Мальпигиево тельце – клубочек артериальных капилляров, охватываемый расширенными стенками канальца, – образует боуменову капсулу. У примитивных форм (акулы, скаты, осетровые) перед капсулой от канальца отходит мерцательная воронка. Мальпигиев клубочек служит аппаратом фильтрации жидких продуктов обмена. В фильтрат попадают как продукты обмена, так и важные для организма вещества. Стенки почечных канальцев пронизаны капиллярами воротных вен и сосудов из боуменовых капсул.
Очищенная кровь возвращается в сосудистую систему почек (почечную вену), а отфильтрованные из крови продукты обмена и мочевина выводятся через каналец в мочеточник. Мочеточники изливаются в мочевой пузырь (мочевой синус), а затем моча выводится наруж 91; у самцов большинства костистых рыб через мочеполовое отверстие позади ануса, а у самок костистых и самцов лососевых, сельдей, щуки некоторых других – через анальное отверстие. У акул и скатов мочеточник открывается в клоаку.
В процессах выделения и водно-солевого обмена кроме почек принимают участие кожа, жаберный эпителий, пищеварительная система (см. ниже).
Жизненная среда рыб – морские и пресные воды – всегда имеет большее или меньшее количество солей, поэтому осморегуляция является важнейшим условием жизнедеятельности рыб.
Осмотическое давление водных животных создается давлением их полостных жидкостей, давлением крови и соков тела. Определяющая роль в этом процессе принадлежит водно-солевому обмену.
Каждая клетка тела имеет оболочку: она полупроницаема, т. е. по-разному проницаема для воды и солей (пропускает воду и солеизбирательно). Водно-солевой обмен клеток определяется в первую очередь осмотическим давлением крови и клеток.
По уровню осмотического давления внутренней среды по отношению к окружающей воде рыбы образуют несколько групп: у миксин полостные жидкости изотоничны окружающей среде; у акул и скатов концентрация солей в жидкостях тела и осмотическое давление немного выше, чем в морской воде, или почти равно ему (достигается за счет разницы солевого состава крови и морской воды и за счет мочевины); у костистых рыб – и морских и пресноводных (как и у более высоко организованных позвоночных) – осмотическое давление внутри тела не равно осмотическому давлению окружающей воды. У пресноводных рыб оно выше, у морских рыб (как и у других позвоночных) ниже, чем в окружающей среде (табл. 2).
Таблица 2
Величина депрессии крови для крупных групп рыб (по Строганову, 1962)
Группа рыб. Депрессия Д°Кровь. Депрессия Д° Внешняя среда. Среднее осмотическое давление, Па. Кровь Среднее осмотическое давление, Па
Внешняя среда.
Костистые: морские. 0,73. 1,90-2,30. 8,9 • 105. 25,1 • 105.
Костистые: пресноводные. 0,52. 0,02-0,03. 6,4 • 105. 0,3 • 105.
Если в организме поддерживается определённый уровень осмотического давления жидкостей тела, то условия жизнедеятельности клеток становятся более стабильными и организм меньше зависит от колебаний внешней среды.
Настоящие рыбы обладают этим свойством – сохранять относительное постоянство осмотического давления крови и лимфы, т. е. внутренней среды; поэтому они относятся к гомойосмотическим организмам (от греч. ‛гомойос‛ – однородный) .
Но у разных групп рыб эта независимость осмотического давления выражается и достигается по-разному,
У морских костистых рыб общее количество солей в крови значительно ниже, чем в морской воде, давление внутренней среды меньше давления внешней, т. е. их кровь гипотонична по отношению к морской воде. Ниже приведены величины депрессии крови рыб (по Строганову, 1962):
Вид рыбы. Депрессия среды Д°.
Морские:
треска балтийская – 0,77
камбала морская – 0,70
скумбрия – 0,73
форель радужная – 0,52
налим – 0,48
Пресноводные:
карп –
0,42
линь – 0,49
щука – 0,52
Проходные:
угорь в море – 0,82
в реке – 0,63
севрюга в море – 0,64
в реке – 0,44
У пресноводных рыб количество солей в крови выше, чем в пресной воде. Давление внутренней среды больше давления внешней, их кровь гипертонична.
Поддержание солевого состава крови и давления ее на нужном уровне обусловливается деятельностью почек, особых клеток стенок почечных канальцев (выделение мочевины), жаберных лепестков (диффузия аммиака, выделение хлоридов), кожных покровов, кишечника, печени.
У морских и пресноводных рыб осморегуляция совершается разными способами (специфическая деятельность почек, различная проницаемость покровов для мочевины, солей и воды, различная деятельность жабр в морской и пресной воде).
У пресноводных рыб (с гипертонической кровью), находящихся в гипотонической среде, разница осмотического давления внутри и вне организма приводит к тому, что вода извне непрерывно поступает внутрь организма – через жабры, кожу и ротовую полость (рис. 23).
Рис. 23. Механизмы осморегуляции у костистых рыб
А – пресноводные; Б – морские (по Строганову, 1962)
Во избежание чрезмерного обводнения, для сохранения водно-солевого состава и уровня осмотического давления возникает необходимость вывода из организма лишней воды и одновременного удержания солей. В связи с этим у пресноводных рыб мощное развитие получают почки. Количество мальпигиевых клубочков и почечных канальцев у них велико; мочи они выделяют гораздо больше, чем близкие морские виды. Данные о количестве мочи, выделяемой рыбами в сутки, представлены ниже (по Строганову, 1962):
Вид рыбы. Количество мочи, мл/кг массы тела
Пресноводные:
карп – 50–120
форель – 60– 106
сом карликовый – 154 – 326
Морские:
бычок –
3–23
морской черт – 18
Проходные:
угорь в пресной воде –
60–150
в море – 2–4
Утрата солей с мочой, экскрементами и через кожу восполняется у пресноводных рыб за счет получения их с пищей благодаря специализированной деятельности жабр (жабры поглощают из пресной воды ионы Na и Сl) и поглощением солей в почечных канальцах.
Морские костистые рыбы (с гипотонической кровью), находящиеся в гипертонической среде, постоянно теряют воду – через кожу, жабры, с мочой, экскрементами. Предотвращение обезвоживания организма и сохранение осмотического давления на нужном уровне (т. е. ниже, чем в морской воде) достигаются тем, что они пьют морскую воду, которая всасывается через стенки желудка и кишечника, а избыток солей выделяется кишечником и жабрами.
Угорь и морской бычок-подкаменщик в морской воде ежедневно пьют 50–200 см3 воды на 1 кг массы тела. В условиях опыта при пре прекращении подачи воды через рот (закрытый пробкой) рыба теряла 12%– 14% массы и на 3–4-й день погибала.
Морские рыбы выделяют очень мало мочи: в почках у них немного мальпигиевых клубочков, у некоторых их нет совсем и есть только почечные канальцы. У них уменьшена проницаемость кожи для солей, жабры выделяют наружу ионы Na и Сl. Железистые клетки стенок канальцев увеличивают выделение мочевины и других прод091;ктов азотисm0;ого обмена.
Таким образом, у непроходных рыб – только морских или только пресноводных – действует какой-нибудь один, специфический для них способ осморегуляции.
Эвригалинные организмы (т. е. выдерживающие значительное колебание солености), в частности проходные рыбы, проводят часть жизни в море, а часть – в пресной воде. При переходе из одной среды в другую, например во время нерестовых миграций, они переносят большие колебания солености.
Это возможно благодаря тому, что проходные рыбы могут переходить с одного способа осморегуляции на другой. В морской воде у них действует такая же система осморегуляции, как у морских рыб, в пресной – как у пресноводных, так что их кровь в морской воде гипотонична, а в пресной – гипертонична.
Их почки, кожа и жабры могут функционировать двояко: почки имеют почечные клубочки с почечными канальцами, как у пресноводных рыб, и только почечные канальцы, как у морских. Жабры снабжены специализированными клетками (так называемые клетки Кейс-Вильмера), способными поглощать и выделять Сl и Na (тогда как у морских или пресноводных рыб они действуют только в одном направлении). Изменяется и количество таких клеток. При переходе из пресной воды в море в жабрах японского угря возрастает количество клеток, выделяющих хлориды. У речной миноги при подъеме из моря в реки количество мочи, выделяемой в течение суток, увеличивается до 45% по сравнению с массой тела.
У некоторых проходных рыб большую роль в регуляции осмотического давления играет слизь, выделяемая кожей.
Передний отдел почки – головная почка – выполняет не выделительную, а кроветворную функцию: в него не заходит воротная вена почек, а в составляющей ее лимфоидной ткани образуются красные и белые кровяные клетки и разрушаются отжившие эритроциты.
Как и селезёнка, почки чутко отражают состояние рыбы, уменьшаясь в объёме при недостатке кислорода в воде и увеличиваясь при замедлении обмена (у карпа – во время зимовки, когда ослабляется деятельность кровеносной системы), в случае острых заболеваний и т. д.
Очень своеобразна дополнительная функция почек у колюшки, строящей для нереста гнездо из кусочков растений: перед нерестом почки увеличиваются, в стенках почечных канальцев вырабатывается большое количество слизи, которая в воде быстро затвердевает и скрепляет гнездо.
НазадОглавлениеДалее
Источник