Где в мозгу находиться центр мочеиспускания

Хранение мочи и периодическое опорожнение мочевого пузыря (МП) зависят от координированной работы гладких и поперечнополосатых мышц, условно разделенных на два структурно функциональных отдела: резервуара (мочевой пузырь) и выпускной системы, состоящей из шейки МП, уретры и наружного сфинктера мочеиспускательного канала. За координированную работу вышеперечисленных структур отвечают сложные нейронные системы управления, расположенные в периферических ганглиях, спинном и головном мозге.
Обеспечение сознательного контроля мочеиспускания требует сложных взаимодействий между вегетативным (симпатика и парасимпатика) и соматическим отделами нервной системы.
Симпатические нервные волокна берут свое начало из боковых рогов грудопоясничного отдела (спинномозговой центр Якобсона) и ганглиях нижнего брыжеечного сплетения и достигают МП в составе подчревного нерва. Симпатические постганглионарные нервы выделяют норадреналин, который активирует β-адренергические рецепторы, ингибирующие сокращение детрузорной мышцы, и α-адренергические рецепторы, возбуждающие мускулатуру уретры и шейки МП. Симпатическое воздействие приводит к расслаблению мышцы, изгоняющей мочу, сокращению мускулатуры уретры и шейки МП и ингибированию интрамуральных ганглиев МП.
Центральный отдел парасимпатической иннервации МП располагается в промежуточных ядрах крестцовых сегментов. Холинергические преганглионарные волокна из промежуточных ядер посылают свои аксоны через тазовые нервы к ганглиозным клеткам тазового сплетения и интрамуральным нейронам в стенке мочевого пузыря. Ганглиозные клетки в свою очередь возбуждают детрузор мочевого пузыря, что приводит к его сокращению с последующим опорожнением. Основными медиаторами парасимпатической нервной системы тут являются ацетилхолин и другие нехолинергические медиаторы. Ацетилхолин действует опосредованно, возбуждая M3-холинорецепторы клеток детрузора. Парасимпатические нервные окончания в нервно-мышечных синапсах и в парасимпатических ганглиях тоже имеют холинорецепторы, возбуждение этих рецепторов на нервных окончаниях может усиливать (через рецепторы М1) или подавлять (через рецепторы М4) высвобождение медиаторов в зависимости от интенсивности нейронного возбуждения. Основной нехолинергический медиатор это АТФ, который активирует внутриклеточную пуринергическую систему через возбуждение P2X рецепторов и тоже способствует сокращению детрузора. Парасимпатические волокна вызывают расслабление ГМК уретры путем высвобождения оксида азота (NO).
Аксоны соматических двигательных нейронов передних рогов крестцовых сегментов S2-S4 (ядро Онуфа) проходят в срамном нерве и иннервируют поперечно-полосатые мышцы наружного уретрального сфинктера. Нейроны более медиально расположенного моторного ядра на том же уровне позвоночника иннервируют мускулатуру тазового дна.
Афферентные пути НМП состоят из цепочек чувствительных нейронов. Первые нейроны, располагающиеся в спинальных ганглиях на уровне S2-S4 и T11-L2, реагируют на пассивное растяжение и активное сокращение мышц МП и передают эту информацию на нейроны второго и третьего порядка. Эти нейроны обеспечивают координированную работу спинальных рефлексов и восходят к вышележащим структурам головного мозга, контролирующих фазы накопления и опорожнения МП. Наиболее важные афферентные волокна от мочевого пузыря идут в составе тазового нерва, в то время как чувствительность от шейки МП и уретры передается по срамным и подчревным нервам. Афферентные волокна этих нервов состоят из миелинизированных (Аδ) и немиелинизированных (С) аксонов. Aδ-волокна передают информацию о наполнении мочевого пузыря. С-волокна нечувствительны к изменению объема мочевого пузыря в физиологических условиях, поэтому они называются «тихими». Они реагируют главным образом на патологические стимулы, такие как химическое раздражение или охлаждение.
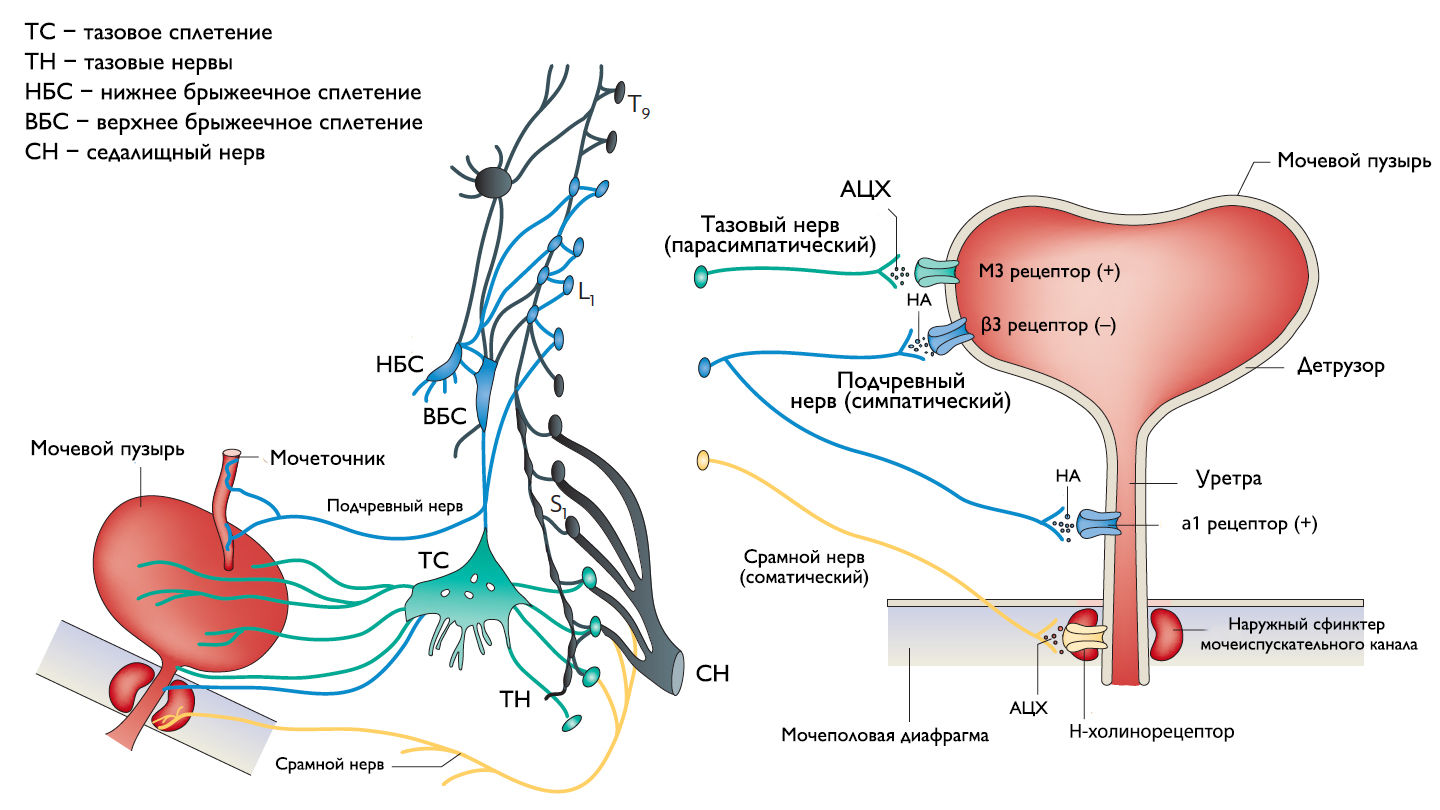
Рисунок 1 | Иннервация нижних мочевыводящих путей.
Специфический и неспецифический восходящий супраспинальный сенсорный путь
Одни спинномозговые промежуточные нейроны посылают восходящие волокна к определенным областям моста и среднего мозга, участвующим в мочеиспускании. Другие промежуточные нейроны передают информацию из нижних мочевыводящих путей в структуры переднего мозга, включая таламус и гипоталамус. Спиноталамический и спиногипоталамический тракты хотя и не играют главную роль в мочеиспускании, но могут включаться в сознательный контроль полноты мочевого пузыря. Чувствительные зоны коры ГМ через спиноталамический тракт информируются о состоянии наполнения МП.
Мостовой центр мочеиспускания (МЦМ) и его нисходящие спинномозговые двигательные пути
Впервые центр управления мочеиспусканием был открыт в дорсальной части моста Баррингтоном в 1925 году и с тех пор называется мостовым центром мочеиспускания или ядром Баррингтона. МЦМ располагается в области покрышки моста. Нейроны МЦМ имеют нисходящие возбуждающие синаптические контакты с клетками парасимпатических преганглионарных мотонейронов, иннервирующих постганглионарные клетки мочевого пузыря. Электрическая и химическая стимуляция МЦМ у крыс и кошек инициирует сокращение мочевого пузыря и расслабляет сфинктер уретры имитируя нормальное мочеиспускание. Цикл рефлекса мочеиспускания состоит из трех фаз, контролируемых различными центральными механизмами: фаза реализации безопасной среды – для начала мочеиспускания человеку необходимо осознание, что окружающие обстановка комфортна; фаза релаксации наружного уретрального сфинктера; и фаза сокращения мышцы, выталкивающей мочу. Процесс нормального мочеиспускания невозможен без какой-либо из этих фаз. МЦМ является командным центром мочеиспускания, который контролирует последовательное переключение фазы расслабления наружного уретрального сфинктера на фазу сокращения детрузорной мышцы.
Мостовой центр удержания мочи (МЦУ) и его нисходящие спинномозговые двигательные пути
Его роль заключается в расслаблении детрузора и сокращении наружного уретрального сфинктера. МЦУ располагается вентролатеральней МЦМ. Синапсы волокон МЦУ возбуждают ядро Онуфа в крестцовых сегментах спинного мозга, повышая таким образом тонус наружного сфинктера уретры. Стимуляция области МЦУ останавливает мочеиспускание, возбуждает мышцы тазового дна и сокращает уретральный сфинктер. Наоборот, двусторонние поражения МЦУ вызывают недержание мочи, чрезмерную детрузорную активность, невозможность хранения мочи и снижение тонуса уретрального сфинктера. На сегодняшний день нет анатомических доказательств связи между МЦУ и МЦМ, и было высказано предположение, что эти центры функционально независимы.
Роль кортикальных областей
Наиболее частые симптомы поражения кортикальных областей ГМ это поллакиурия и ургентное недержание мочи. Поэтому Andrew и Nathan выдвинули гипотезу, что отсоединение лобной или передней поясной извилины от гипоталамуса приводит к непроизвольному началу мочеиспускания [3]. Действительно, префронтальная кора головного мозга человека и передняя поясная извилина активируются во время мочеиспускания [4].
Мозжечок и базальные ганглии
Существует ряд исследований о том, что мозжечок и базальные ганглии оказывают в основном ингибирующее действие на мочевой пузырь. Мозжечковая патология приводит к увеличению частоты мочеиспускания и ургентному недержанию мочи. Симптомы гиперактивного мочевого пузыря также встречаются при болезни Паркинсона. Поскольку нет прямых связей этих областей с МЦМ, ингибирующее влияние, вероятно, косвенное через структуры переднего и среднего мозга.
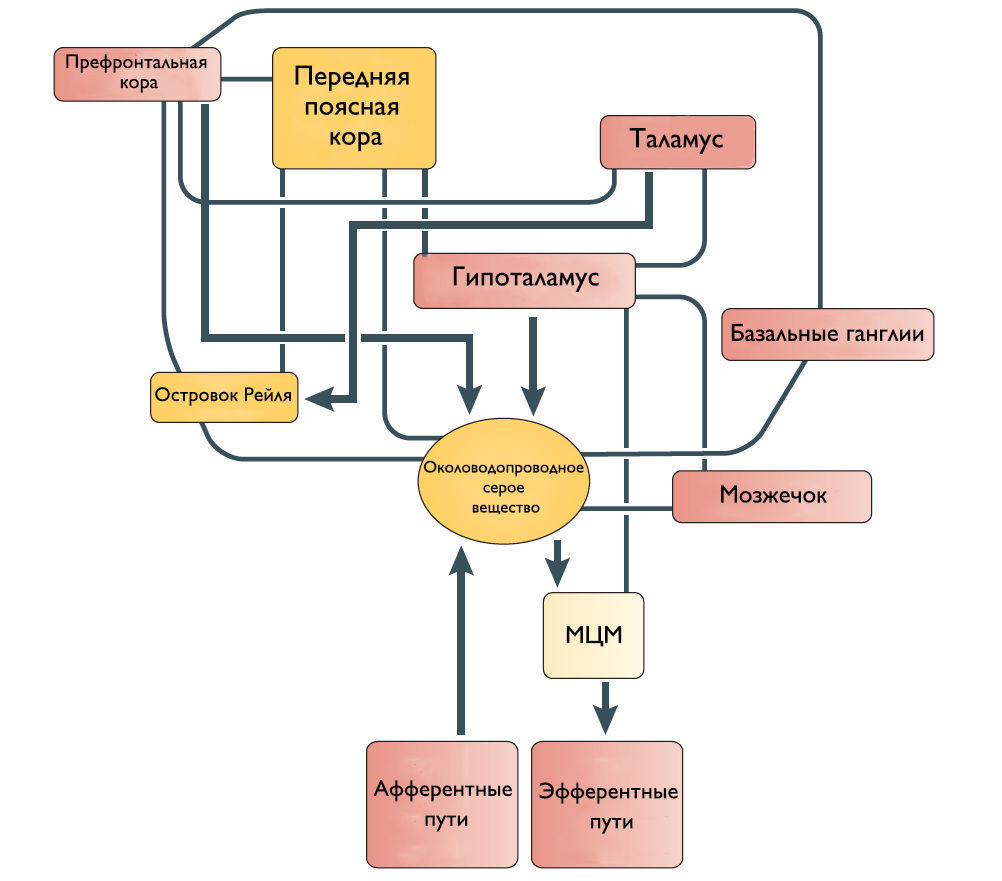
Рисунок 2 | Предположительное схематическое изображение связей между различными структурами переднего мозга и ствола мозга, которые участвуют в контроле мочеиспускания.
- Liao L., Madersbacher H. (ed.). Neurourology: Theory and Practice. – Springer, 2019.
- Clare J. Fowler et al. The neural control of micturition. – Nature Reviews | Neuroscience, volume 9. – June 2008.
- Andrew J, Nathan PW. Lesions of the anterior frontal lobes and disturbances of micturition and defecation. Brain. 1964;87:233-62.
- Griffiths, Derek J. “Use of al imaging to monitor central control of voiding in humans.” Urinary Tract. Springer, Berlin, Heidelberg, 2011. 81-97.
Нашли опечатку? Выделите фрагмент и нажмите Ctrl+Enter.
Источник
«Нет в жизни большего счастья, чем вовремя освобожденный мочевой пузырь» (Овидий)
«Хорошее мочеиспускание – это единственное удовольствие, которое можно получить, не испытывая потом угрызений совести» (И. Кант)
Ежечасно в мочевой пузырь у здорового взрослого человека поступает примерно до 50 мл мочи, что постепенно по мере наполнения пузыря повышает в нем давление. При достижении объема около 400 мл возникает чувство наполнения пузыря. Рефлекс мочеиспускания может быть реализован при количестве мочи от 300 до 500 мл (в зависимости от антропометрических показателей индивидуума). Но прежде чем переходить к рассмотрению процесса мочеиспускания и его регуляции необходимо ознакомится с субстратом этого процесса (с анатомической точки зрения), т.е. с мочевым пузырем, а точнее с его сфинктерами и детрузором.
Детрузор мочевого пузыря (от лат. «detrudere» – выталкивать) – это мышечная оболочка (мочевого пузыря), состоящая из трех взаимно переплетающихся слоев, которые образуют единую, изгоняющую мочу мышцу – детрузор (m. detrusor urinae). Таким образом, сокращение детрузора приводит к мочеиспусканию. Наружный слой детрузора состоит из продольных волокон, средний – из циркулярных и внутренний – из продольных и поперечных. Наиболее развит средний слой, который в области внутреннего отверстия уретры образует сфинктер шейки мочевого пузыря или внутренний сфинктер (! обратите внимание – анатомическая общность предполагает и общую иннервацию детрузора и внутреннего сфинктера мочевого пузыря, т.е. при мочеиспускании происходит одновременное – рефлекторное, – расслабление внутреннего сфинктера и сокращение мочевого пузыря). Следует обратить внимание на то, что мышцы, составляющие внутренний сфинктер мочевого пузыря и m.detrusor urinae, состоят из гладких мышечных волокон, получающих вегетативную иннервацию, и поэтому не подчиняются сознанию. Наружный сфинктер находится он на уровне тазового дна и состоит из поперечнополосатой мускулатуры, иннервируемой соматическими нервами и, как следствие, подчиняется сознанию. Такой осознанный контроль способен подавить непроизвольную попытку опорожнить мочевой пузырь, т.е. (в норме) моча не выходит, пока человек «сознательно не решит открыть сфинктер».
Весьма нередко в современной научной и учебной литературе, к сожалению, приходится сталкиваться с утверждением о наличии 2-х (внутреннего и наружного) сфинктеров мочевого пузыря. У мочевого пузыря нет ни одного сфинктера. То, что именуют внутренним «гладкомышечным» сфинктером таковым не является, поскольку не содержит циркулярных мышечных волокон, присущих сфинктерам. То, что расположено вокруг внутреннего отверстия уретры и ее проксимального отдела – это комплекс анатомических образований: язычок пузыря «uvula vesicae» – кавернозноподобное образование пузырно-уретрального сегмента, петля детрузора, пучки продольных гладкомышечных волокон, переходящих от детрузора к уретре и поперечные гладкомышечные пучки латеральных отделов проксимальной уретры. Кровенаполнение «язычка» способствует удержанию мочи в пузыре, петля фиксирует пластинку основания. Продольные волокна при сокращении укорачивают проксимальный отдел уретры, способствуя раскрытию ее внутреннего отверстия перед мочеиспусканием, а поперечные – обусловливают смыкание передней и задней стенок проксимального отдела уретры для удержания мочи. «Наружный» сфинктер, действительно содержащий циркулярные гладкомышечные волокна, не относится к мочевому пузырю, а, как известно, является сфинктером уретры.
источник «Нарушения функции мочевого пузыря (лекция)» Борисов В.В. Кафедра нефрологии и гемодиализа ФППО врачей Первого Московского государственного медицинского университета им. И.М. Сеченова, Москва (журнал «Вестник урологии» №1 – 2014) [читать]
цитата из клинической лекции «Особенности деятельности мочевого пузыря» В.В. Борисов:
« … Особое место в обеспечении функции мочевого пузыря занимает структура мелких внутристеночных сосудов, которые имеют спиральную форму. Именно она позволяет сохранять необходимый постоянный просвет в условиях значительного растяжения стенки. При этом растягиваются спирали, а просвет артериального сосуда остается неизменным. Не менее важное значение в обеспечении функции системы мочевыводящих путей в целом и мочевого пузыря в частности имеют кавернозноподобные сосудистые образования, открытые в стенке мочеточника и мочевого пузыря Ю.А. Пытелем в середине прошлого века и подтвержденные дальнейшими исследованиями морфологов школы академика В.В. Куприянова. По своей структуре они напоминают кавернозную ткань полового члена, в которой как в губке может депонироваться кровь, значительно увеличивая объем этого образования. Внезапное переполнение такого образования кровью способствует сокращениям окружающих гладкомышечных структур и осуществлению быстрого и эффективного перекрытия просвета полого органа. Такие образования были описаны в области лоханочно-мочеточникового, мочеточниково-пузырного и пузырно-уретрального сегментов мочевыводящих путей. Для мочевого пузыря кавернозноподобные образования в области мочеточникового устья являются одним из антирефлюксных механизмов при мочеиспускании, а в области шейки мочевого пузыря – одним из механизмов удержания мочи в пузыре в фазу наполнения …» [читать лекцию полностью]
В сущности детрузор – целостная мышца, единый функциональный синцитий гладкомышечных клеток и волокон, ориентированных спирально во взаимно перпендикулярных плоскостях, волокон, которые переходят из внутренних слоев в средние и наружные и наоборот. Именно эта особенность строения позволяет детрузору работать содружественно и на активное расширение в фазу наполнения, и на активное сокращение при опорожнении мочевого пузыря.
Деятельность мочевого пузыря многогранна и включает накопление и удержание мочи, эвакуацию мочи по уретре наружу (т.е. мочеиспускание), а также, что не менее важно, облегчение поступлений порций мочи из терминальных отделов мочеточников и предотвращение обратного поступления мочи из мочевого пузыря в мочеточники.
Нейрогенные регуляторные механизмы деятельности мочевого пузыря сложны, являются элементами вегетативной нервной системы и имеют представительство в коре, лимбической системе, таламусе, гипоталамусе, ретикулярной формации, а также связаны с мозжечком. Проводящими путями они связаны с центром мочеиспускания в нижне-поясничном и крестцовом отделах спинного мозга. Сфинктер уретры с помощью срамного (син.: полового) нерва получает не только вегетативную, но и соматическую иннервацию, определяющую произвольное мочеиспускание.
Высшим центром регуляции всей системы, управляющей мочеиспусканием является головной мозг, в котором центр мочеиспускания последнего располагается в парацентральной дольке лобной доли (по соседству с центром стопы). Основной функцией центра мочеиспускания, включающего лобную долю, является (! произвольное, осознанное) тоническое торможение сокращения детрузора до наиболее подходящего благоприятного момента для опорожнения мочевого пузыря.
[читать] статью «Роль головного мозга в регуляции процесса мочеиспускания» В.Б. Бердичевский, А.А. Суфианов, В.Г. Елишев, Д.А. Барашин, Клиника урологии ГБОУ ВПО «Тюменская государственная медицинская академия» Минздрава России (журнал «Андрология и генитальная хирургия» №1, 2014)
Следующим центром в системе нервного контроля над мочеиспусканием является центр, расположенный в мосте. Также он имеет название ядра Баррингтона или Nucleus Locus Coerulus (ядро голубоватого места). Центр локализуется в вентральной части серого веществе, расположенного вокруг водопровода. В заднем отделе покрышки моста выделяют две взаимодействующие области: М-зона (зона опорожнения) и L-зона (зона накопления). Центр мочеиспускания моста играет роль основного релейного переключателя афферентных и эфферентных импульсов между головным мозгом и нижними мочевыми путями (мочевой пузырь, уретра). Он также координирует последовательное расслабление сфинктера уретры и сокращение детрузора при мочеиспускании.
Низшие центры (парасимпатический и симпатический), которые осуществляют (! непроизвольно, неосознанно) акт мочеиспускания, расположены в спинном мозге. Кроме того, в спинном мозге расположены проводящие нервные волокна, соединяющие высшие (парацентральные дольки, ядра Баррингтона) и низшие (спинномозговые центры) мочеиспускания. Парасимпатический центр мочеиспускания расположен в крестцовом (сакральном) отделе спинного мозга (в сегментах S2 – S4). Симпатический центр мочеиспускания расположен в грудо-поясничном отделе спинного мозга (в сегментах Т9-10 – L2-3). Классическая концепция деятельности мочевого пузыря в целом предполагает, что фаза наполнения (расслабление детрузора и сокращение, закрытие сфинктеров) является симпатической, а мочеиспускание (сокращение детрузора и расслабление, раскрытие сфинктеров) реализуется парасимпатическими структурами.
Соматические нервы. Как было указано выше, в спинном мозге расположены проводящие нервные волокна, соединяющие высшие и низшие, спинномозговые, центры мочеиспускания (в сегментах S2-4), что позволяет осуществлять произвольный нисходящий контроль над актом мочеиспускания. Это «соединение» осуществляется пирамидными (двигательными) путями. От спинного мозга до мочевого пузыря дальнейшее соединение осуществляется соматическими (половыми) нервами, основной точкой приложения которых является наружный сфинктер; причем этот сфинктер может сокращаться произвольно, но расслабляется он рефлекторно вместе с открытием внутреннего сфинктера при начале мочеиспускания. В основном наружный сфинктер обеспечивает удержание мочи (произвольное, осознанное) при повышении давления в мочевом пузыре.
Чувствительная иннервация мочевого пузыря. Афферентные (идущие от периферии к центру) волокна начинаются в рецепторах, расположенных в стенке мочевого пузыря и реагирующих на растяжение. Наполнение мочевого пузыря рефлекторно повышает тонус мышц стенки мочевого пузыря и внутреннего сфинктера, которые иннервируются нейронами крестцовых сегментов (S2-4) и внутренностными тазовыми нервами. Усиление давления на стенку мочевого пузыря воспринимается осознанно, поскольку часть афферентных импульсов по задним канатикам спинного мозга устремляются к центру мочеиспускания в стволе мозга, который размещается в ретикулярной формации вблизи от голубоватого места. От центра мочеиспускания импульсы следуют в парацентральную дольку на медиальной поверхности больших полушарий и к другим областям мозга.
Предполагается, что в процессе эволюции первоначально сформировавшаяся нервная система разделилась на анимальную и вегетативную нервную систему. Анимальная нервная система, связанная с деятельностью органов чувств и произвольной скелетной мускулатуры, обеспечивала приспособление организма к действию факторов окружающей среды. Ее функции контролируются сознанием. Вегетативная нервная система, регулируя деятельность внутренних органов, обеспечивала сохранение постоянства внутренней среды организма. В ответ на негативное влияние внешних факторов она, мобилизуя адаптационно-компенсаторные механизмы организма, способствовала выполнению функций анимальной нервной системы. Деятельность вегетативной нервной системы осуществлялась без участия сознания. Симпатическая часть автономной нервной системы брала на себя адаптацию организма к условиям внешней среды. Парасимпатическая часть автономной нервной системы способствовала сохранению постоянства внутренней среды организма. Метасимпатическая часть автономной нервной системы обеспечивала врожденный автоматизм органа и являлась эволюционно самой древней четью вегетативной нервной системы. Сфера ее иннервации ограничена и охватывает сугубо полый орган. Эта автономия интрамуральных ганглиев, имея полный набор необходимых для самостоятельной рефлекторной деятельности звеньев – сенсорного, ассоциативного, эффекторного, представляет собой как бы собственный «мозг» органа. В эксперименте показано, что, обладая значительной независимостью от центрального и периферического регулирования, метасимпатическая нервная система способна осуществлять адекватную рефлекторную деятельность органа при полной его денервации. Так, свежеэкстирпированный мочевой пузырь животного при достаточном наполнении через уретру теплым солевым раствором, способен к спонтанному опорожнению. Не все ученые готовы признать выделение метасимпатической нервной системы в самостоятельный раздел нервной системы, считая ее частью парасимпатической иннервации мочевого пузыря. Однако наличие у органа существенных автономных свойств никто не отрицает.
Весь механизм накопления и опорожнения мочевого пузыря схематично выглядит следующим образом. В процессе физиологического обеспечения работы нижних мочевых путей организм человека создает и поддерживает определенный тонус поперечно-полосатых мышц передней стенки живота и промежности. В этих комфортных условиях, исходя из наличия автономных (непроизвольных, неконтролируемых сознанием) свойств, мочевой пузырь медленно накапливает мочу в расслабленный резервуар детрузора. Сомато-висцеральный рефлекс обеспечивает процесс удержания поступившей на хранение мочи посредством повышенного тонуса внутреннего и наружного сфинктеров мочевого пузыря, а также исходным тонусом мышц промежности. Физиологический тонус поперечно-полосатых мышц тела человека указывает на адекватную работу головного мозга, в рамках сознательного контроля за функцией мочевого пузыря, в условиях адаптации организма человека к внешним факторам пребывания. Центральная нервная система одновременно оказывает корректирующее воздействие на работу вегетативной нервной системы, обеспечивающей поддержание гомеостаза, в том числе и резервуарных функций мочевого пузыря. Физиологически преобладает симпатикотония мочевого пузыря. Детрузор расслаблен. Его размер медленно адаптируется под объем поступающей мочи. При этом ведущей функцией симпатической нервной системы является нивелирование внутрипузырного давления путем синхронного увеличения емкости мочевого пузыря. Парасимпатическая нервная система находится в угнетенном состоянии. Она не посылает импульсы на сокращение детрузора и расслабление внутреннего сфинктера. Все системы, регулирующие накопление и удержание мочи, находятся в состоянии функционального равновесия. Мочевой пузырь наполняется мочой до физиологически приемлемого уровня. Нервные импульсы об этом по латеральным канатикам спинного мозга поступают в парацентральные дольки больших полушарий, часть импульсов переходит на противоположную сторону. Осознанная регуляция мочеиспускания осуществляется благодаря нервной импульсации от двигательной зоны коры больших полушарий к мотонейронам передних рогов сегментов S2-4. Головной мозг с целью инициации акта мочеиспускания дает команду мышцам брюшного пресса на сокращение, и одновременно мышцам наружного сфинктера мочевого пузыря для беспрепятственного обеспечения этого процесса. Реализуется сомато-висцеральный рефлекс. Эта импульсация одновременное оказывает пусковое воздействие на метасимпатическую часть нервной системы мочевого пузыря и корректирующее – на другие вегетативные центры. Симпатическое доминирование угасает, и мочевой пузырь переходит под влияние парасиматической иннеравции. Наступает фаза парасимпатикотонии мочевого пузыря. Под влиянием ацетилхолина (медиатора парасимпатической нервной системы) детрузор сокращается, внутренний сфинктер мочевого пузыря расслабляется. Все происходит быстро, синхронно, и весь объем накопленной мочи покидает мочевой пузырь. Головной мозг информируется органами внешнего контроля (слух, зрение, тактильные ощущения) о завершении акта мочеиспускания. Висцеро-соматический рефлекс побуждает к сокращению мышц промежности и расслаблению передней брюшной стенки, с последующим переводом их в режим физиологического тонуса. Одновременно автономные функции мочевого пузыря ставятся под охрану вегетативных центров, сопровождающих новый процесс наполнения мочевого пузыря в рамках поддержания гомеостаза организма человека.
В жизненном пространстве человека доминирует система удержания мочи, преимущественно регулируемая симпатическим отделом вегетативной нервной системы. Осознанное ощущение полноты мочевого пузыря опосредовано растяжением стенки органа возрастающим объемом мочи в фазу наполнения. При этом чувствительные импульсы от рецепторов, расположенных в его стенке, по тазовому нерву поступают в крестцовый отдел спинного мозга. Далее они направляются по передним и задним столбам спинного мозга в центры мочеиспускания, расположенные в области моста и коры головного мозга. Головной мозг снабжен органами внешнего контроля, которые оценивают сложившуюся жизненно важную ситуацию. Если на данный отрезок времени для конкретного индивидуума существует подходящая обстановка, то головной мозг, ощущающий позыв на мочеиспускание, конкретными действиями инициирует начало акта мочеиспускания. Одновременно плавно напрягаются мышцы живота, иннервируемые межреберными нервами, и расслабляются мышцы промежности за счет эфферентных соматических импульсов, достигающих мишени по половому нерву. Это осознанный и управляемый этап мочеиспускания. Далее этот соматический импульс подавляет симпатическое доминирование над мочевым пузырем, обеспечивающим медленное накопление мочи, и активизирует парасимпатическое влияние на орган, через эфферентные пути тазового нерва для быстрого и исчерпывающего опорожнения последнего.
Отсутствие комфортных условий для акта мочеиспускания заставляет человека волевым решением подавить соматическую импульсацию в виде позыва на мочеиспускание и передать команду симпатической иннервации продолжить процесс накопления мочи инициируемого медиатором норадреналином. Следующий позыв на мочеиспускание также может совпасть с отсутствием надлежащих условий. И снова головной мозг подавляет реакции спинного мозга, направленные на исполнение процесса избавления мочевого пузыря от нарастающего объема мочи. Позыв вновь перестает быть актуальным для поведения человека. Третий позыв на мочеиспускание тревожит головной мозг на пределе объемных возможностей мочевого пузыря. Условий для мочеиспускания по-прежнему нет. Сознание и воспитание не позволяют исполнению востребованного физиологического акта. Однако человек ощущает, что более не может сопротивляться нарастающему давлению мочи на управляемые мышцы промежности, уретры, и мощная струя как бы исподволь покидает мочевой тракт. Это результат повелительного позыва на мочеиспускание, который, игнорируя запретные усилия сознания и запрещающее координирующее влияние вегетативной нервной системы, побуждает автономную метасимпатическую нервную систему срочно и эффективно избавить мочевой пузырь от «угрожающего для жизни» объема мочи. И только легкий румянец стыда укажет на вынужденное неповиновение мочевого пузыря центральной и вегетативной вертикали управления нервной системы.
Источник