Функции промежуточного мозга мочеиспускание

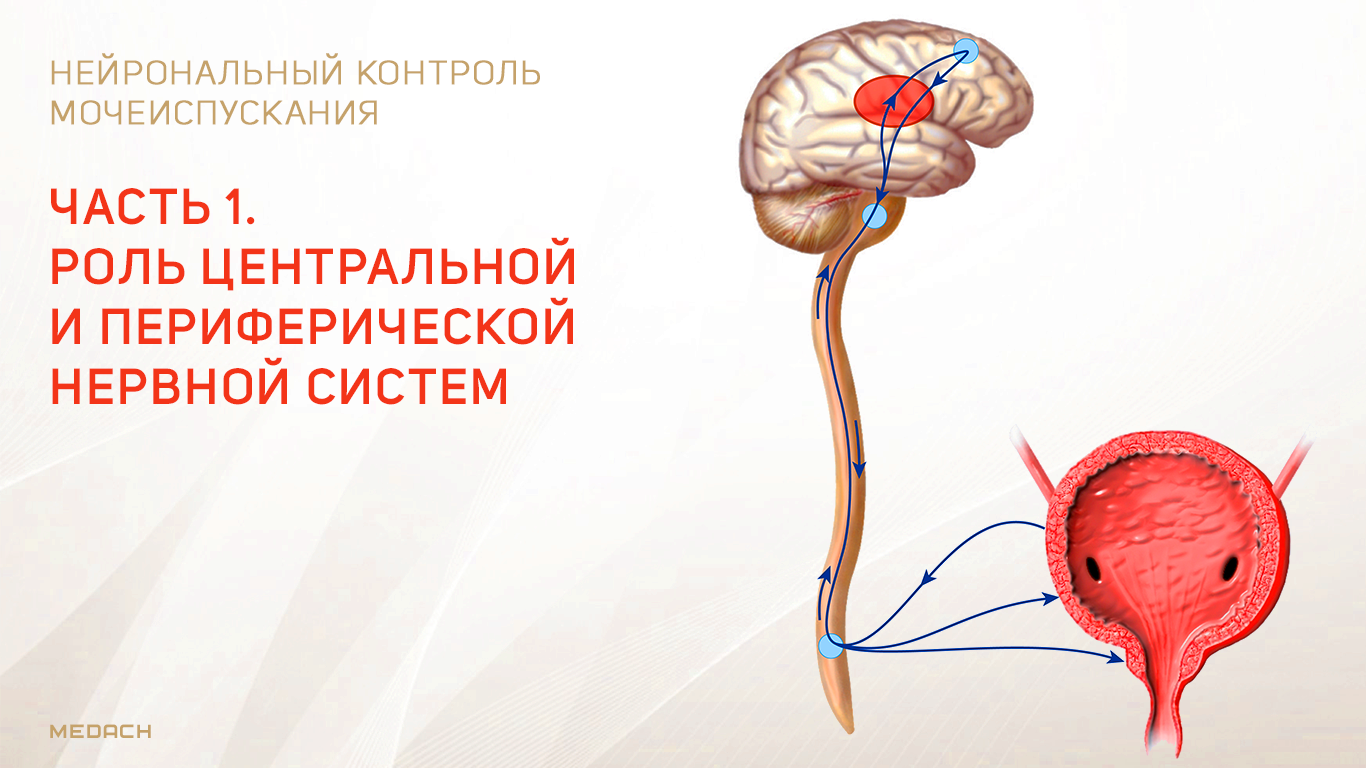
Хранение мочи и периодическое опорожнение мочевого пузыря (МП) зависят от координированной работы гладких и поперечнополосатых мышц, условно разделенных на два структурно функциональных отдела: резервуара (мочевой пузырь) и выпускной системы, состоящей из шейки МП, уретры и наружного сфинктера мочеиспускательного канала. За координированную работу вышеперечисленных структур отвечают сложные нейронные системы управления, расположенные в периферических ганглиях, спинном и головном мозге.
Обеспечение сознательного контроля мочеиспускания требует сложных взаимодействий между вегетативным (симпатика и парасимпатика) и соматическим отделами нервной системы.
Симпатические нервные волокна берут свое начало из боковых рогов грудопоясничного отдела (спинномозговой центр Якобсона) и ганглиях нижнего брыжеечного сплетения и достигают МП в составе подчревного нерва. Симпатические постганглионарные нервы выделяют норадреналин, который активирует β-адренергические рецепторы, ингибирующие сокращение детрузорной мышцы, и α-адренергические рецепторы, возбуждающие мускулатуру уретры и шейки МП. Симпатическое воздействие приводит к расслаблению мышцы, изгоняющей мочу, сокращению мускулатуры уретры и шейки МП и ингибированию интрамуральных ганглиев МП.
Центральный отдел парасимпатической иннервации МП располагается в промежуточных ядрах крестцовых сегментов. Холинергические преганглионарные волокна из промежуточных ядер посылают свои аксоны через тазовые нервы к ганглиозным клеткам тазового сплетения и интрамуральным нейронам в стенке мочевого пузыря. Ганглиозные клетки в свою очередь возбуждают детрузор мочевого пузыря, что приводит к его сокращению с последующим опорожнением. Основными медиаторами парасимпатической нервной системы тут являются ацетилхолин и другие нехолинергические медиаторы. Ацетилхолин действует опосредованно, возбуждая M3-холинорецепторы клеток детрузора. Парасимпатические нервные окончания в нервно-мышечных синапсах и в парасимпатических ганглиях тоже имеют холинорецепторы, возбуждение этих рецепторов на нервных окончаниях может усиливать (через рецепторы М1) или подавлять (через рецепторы М4) высвобождение медиаторов в зависимости от интенсивности нейронного возбуждения. Основной нехолинергический медиатор это АТФ, который активирует внутриклеточную пуринергическую систему через возбуждение P2X рецепторов и тоже способствует сокращению детрузора. Парасимпатические волокна вызывают расслабление ГМК уретры путем высвобождения оксида азота (NO).
Аксоны соматических двигательных нейронов передних рогов крестцовых сегментов S2-S4 (ядро Онуфа) проходят в срамном нерве и иннервируют поперечно-полосатые мышцы наружного уретрального сфинктера. Нейроны более медиально расположенного моторного ядра на том же уровне позвоночника иннервируют мускулатуру тазового дна.
Афферентные пути НМП состоят из цепочек чувствительных нейронов. Первые нейроны, располагающиеся в спинальных ганглиях на уровне S2-S4 и T11-L2, реагируют на пассивное растяжение и активное сокращение мышц МП и передают эту информацию на нейроны второго и третьего порядка. Эти нейроны обеспечивают координированную работу спинальных рефлексов и восходят к вышележащим структурам головного мозга, контролирующих фазы накопления и опорожнения МП. Наиболее важные афферентные волокна от мочевого пузыря идут в составе тазового нерва, в то время как чувствительность от шейки МП и уретры передается по срамным и подчревным нервам. Афферентные волокна этих нервов состоят из миелинизированных (Аδ) и немиелинизированных (С) аксонов. Aδ-волокна передают информацию о наполнении мочевого пузыря. С-волокна нечувствительны к изменению объема мочевого пузыря в физиологических условиях, поэтому они называются «тихими». Они реагируют главным образом на патологические стимулы, такие как химическое раздражение или охлаждение.
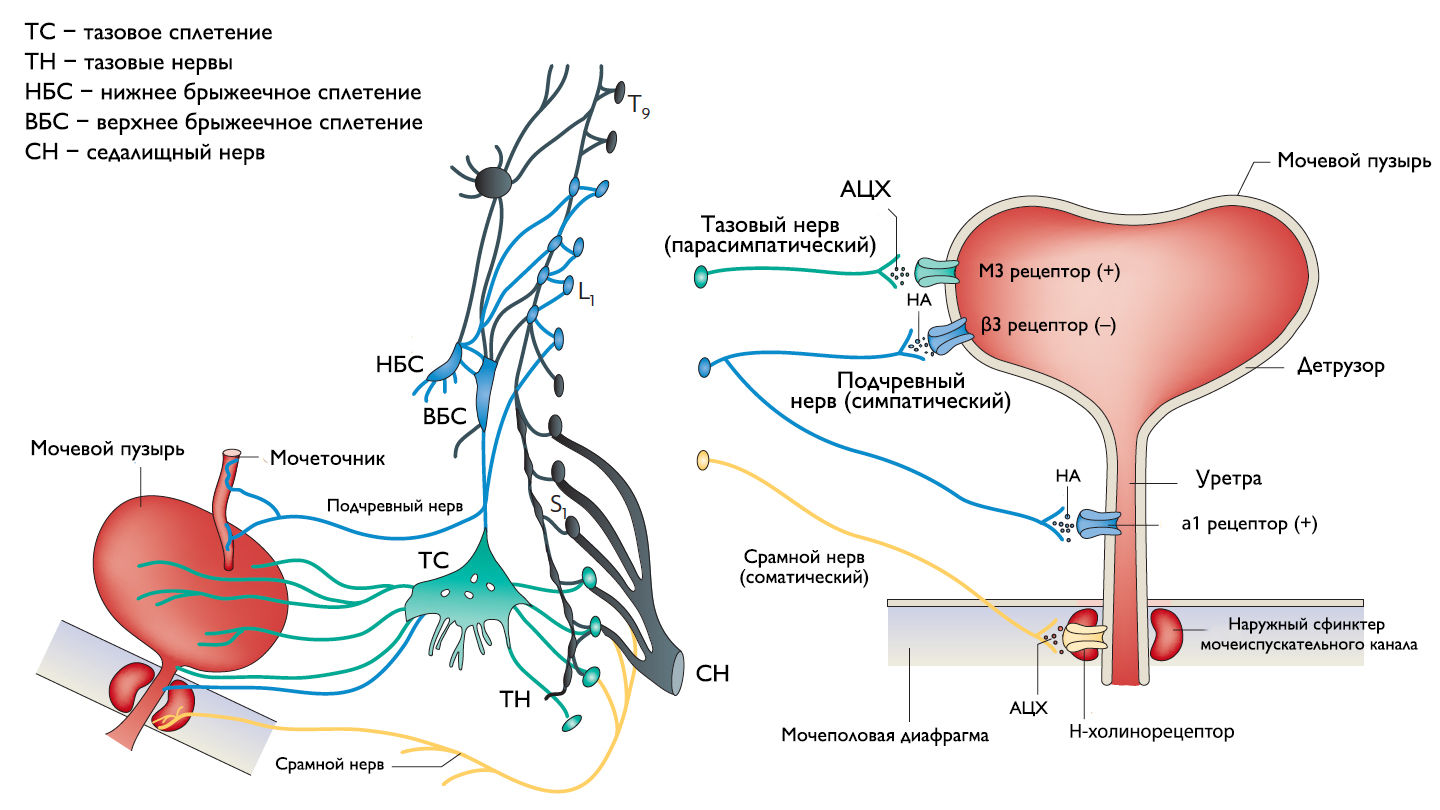
Рисунок 1 | Иннервация нижних мочевыводящих путей.
Специфический и неспецифический восходящий супраспинальный сенсорный путь
Одни спинномозговые промежуточные нейроны посылают восходящие волокна к определенным областям моста и среднего мозга, участвующим в мочеиспускании. Другие промежуточные нейроны передают информацию из нижних мочевыводящих путей в структуры переднего мозга, включая таламус и гипоталамус. Спиноталамический и спиногипоталамический тракты хотя и не играют главную роль в мочеиспускании, но могут включаться в сознательный контроль полноты мочевого пузыря. Чувствительные зоны коры ГМ через спиноталамический тракт информируются о состоянии наполнения МП.
Мостовой центр мочеиспускания (МЦМ) и его нисходящие спинномозговые двигательные пути
Впервые центр управления мочеиспусканием был открыт в дорсальной части моста Баррингтоном в 1925 году и с тех пор называется мостовым центром мочеиспускания или ядром Баррингтона. МЦМ располагается в области покрышки моста. Нейроны МЦМ имеют нисходящие возбуждающие синаптические контакты с клетками парасимпатических преганглионарных мотонейронов, иннервирующих постганглионарные клетки мочевого пузыря. Электрическая и химическая стимуляция МЦМ у крыс и кошек инициирует сокращение мочевого пузыря и расслабляет сфинктер уретры имитируя нормальное мочеиспускание. Цикл рефлекса мочеиспускания состоит из трех фаз, контролируемых различными центральными механизмами: фаза реализации безопасной среды – для начала мочеиспускания человеку необходимо осознание, что окружающие обстановка комфортна; фаза релаксации наружного уретрального сфинктера; и фаза сокращения мышцы, выталкивающей мочу. Процесс нормального мочеиспускания невозможен без какой-либо из этих фаз. МЦМ является командным центром мочеиспускания, который контролирует последовательное переключение фазы расслабления наружного уретрального сфинктера на фазу сокращения детрузорной мышцы.
Мостовой центр удержания мочи (МЦУ) и его нисходящие спинномозговые двигательные пути
Его роль заключается в расслаблении детрузора и сокращении наружного уретрального сфинктера. МЦУ располагается вентролатеральней МЦМ. Синапсы волокон МЦУ возбуждают ядро Онуфа в крестцовых сегментах спинного мозга, повышая таким образом тонус наружного сфинктера уретры. Стимуляция области МЦУ останавливает мочеиспускание, возбуждает мышцы тазового дна и сокращает уретральный сфинктер. Наоборот, двусторонние поражения МЦУ вызывают недержание мочи, чрезмерную детрузорную активность, невозможность хранения мочи и снижение тонуса уретрального сфинктера. На сегодняшний день нет анатомических доказательств связи между МЦУ и МЦМ, и было высказано предположение, что эти центры функционально независимы.
Роль кортикальных областей
Наиболее частые симптомы поражения кортикальных областей ГМ это поллакиурия и ургентное недержание мочи. Поэтому Andrew и Nathan выдвинули гипотезу, что отсоединение лобной или передней поясной извилины от гипоталамуса приводит к непроизвольному началу мочеиспускания [3]. Действительно, префронтальная кора головного мозга человека и передняя поясная извилина активируются во время мочеиспускания [4].
Мозжечок и базальные ганглии
Существует ряд исследований о том, что мозжечок и базальные ганглии оказывают в основном ингибирующее действие на мочевой пузырь. Мозжечковая патология приводит к увеличению частоты мочеиспускания и ургентному недержанию мочи. Симптомы гиперактивного мочевого пузыря также встречаются при болезни Паркинсона. Поскольку нет прямых связей этих областей с МЦМ, ингибирующее влияние, вероятно, косвенное через структуры переднего и среднего мозга.
Рисунок 2 | Предположительное схематическое изображение связей между различными структурами переднего мозга и ствола мозга, которые участвуют в контроле мочеиспускания.
- Liao L., Madersbacher H. (ed.). Neurourology: Theory and Practice. – Springer, 2019.
- Clare J. Fowler et al. The neural control of micturition. – Nature Reviews | Neuroscience, volume 9. – June 2008.
- Andrew J, Nathan PW. Lesions of the anterior frontal lobes and disturbances of micturition and defecation. Brain. 1964;87:233-62.
- Griffiths, Derek J. “Use of al imaging to monitor central control of voiding in humans.” Urinary Tract. Springer, Berlin, Heidelberg, 2011. 81-97.
Нашли опечатку? Выделите фрагмент и нажмите Ctrl+Enter.
Источник
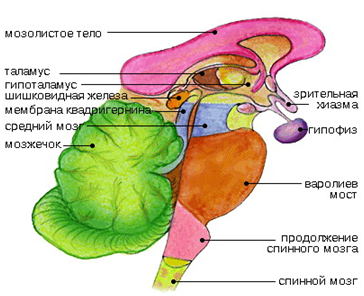 Промежуточный мозг включает в себя таламус, эпиталамус (эпифиз) и гипоталамус и образует стенки III желудочка. Промежуточный мозг регулирует сложные двигательные рефлексы, координирует работу внутренних органов и осуществляет гуморальную регуляцию (обмен веществ, потребление воды и пищи, поддержание температуры тела). Вместе с большими полушариями промежуточный мозг участвует в организации всех сложных форм поведения, регуляции вегетативных реакций, то есть он интегрирует сенсорные, двигательные и вегетативные функции, обеспечивая деятельность организма как единого целого.
Промежуточный мозг включает в себя таламус, эпиталамус (эпифиз) и гипоталамус и образует стенки III желудочка. Промежуточный мозг регулирует сложные двигательные рефлексы, координирует работу внутренних органов и осуществляет гуморальную регуляцию (обмен веществ, потребление воды и пищи, поддержание температуры тела). Вместе с большими полушариями промежуточный мозг участвует в организации всех сложных форм поведения, регуляции вегетативных реакций, то есть он интегрирует сенсорные, двигательные и вегетативные функции, обеспечивая деятельность организма как единого целого.
Таламус (зрительный бугор) представляет собой комплекс ядер в промежуточном мозге (у человека примерно 60 ядер), образующих множество прямых и обратных связей с корой больших полушарий. В таламусе осуществляется анализ афферентных сигналов практически от всех чувствительных рецепторов (кроме обонятельных), организация интегративных процессов, необходимых для регуляции функционального состояния и высшей нервной деятельности.
Ядра таламуса делят на две группы – специфические и неспецифические. Все ядра таламуса в разной степени обладают тремя общими функциями – переключающей, интегративной и модулирующей.
Специфические ядра таламуса:
1) Переключающие ядра:
1а) сенсорные – передают афферентную (чувствительную) информацию в сенсорные зоны коры;
1б) несенсорные – переключают в кору несенсорную импульсацию из разных отделов головного мозга (например, лимбические ядра таламуса).
2) Ассоциативные ядра – принимают импульсацию от других ядер таламуса. Благодаря их деятельности осуществляется объединение деятельности таламических ядер и различных зон ассоциативной коры.
Неспецифические ядра действуют как объединяющие посредники между стволом мозга и мозжечком, с одной стороны, и новой корой, лимбической системой и базальными ганглиями, с другой стороны, объединяя их в единую функциональную систему. Они обеспечивают модулирование, плавную настройку функционирования ЦНС. По своему функциональному значению они сходны с ретикулярной формацией. Но если ретикулярная формация осуществляет длительную и медленную активацию коры больших полушарий, то неспецифические ядра таламуса – быструю и кратковременную активацию.
Гипоталамус является вентральной частью промежуточного мозга. Макроскопически он включает в себя преоптическую область и область перекреста зрительных нервов, серый бугор и воронку, сосцевидные тела. Микроскопически в гипоталамусе выделяют около 50 пар ядер, которые топографически объединяют в 5 групп. Ядра гипоталамуса имеют мощное кровоснабжение.
Функции гипоталамуса:
- В ядрах гипоталамуса локализуются центры, участвующие в вегетативной регуляции, а также нейроны, осуществляющие секрецию нейрогормонов.
- Центр гомеостаза. Нейроны гипоталамуса могут реагировать на изменения температуры крови, электролитного состава, осмотического давления плазмы, количества и состав гормонов крови (благодаря повышенной проницаемости гематоэнцефалического барьера для многих веществ в области гипоталамуса).
- Центры терморегуляции. В ядрах передней группы – центр физической терморегуляции (регуляция теплоотдачи), в ядрах задней группы – центр химической терморегуляции (регуляция теплопродукции).
- Центры регуляции водного и солевого обмена. Среди нейронов паравентрикулярного и супраоптического ядер есть нейроны, продуцирующие антидиуретический гормон, а в латеральном гипоталамическом ядре – центр жажды, обеспечивающий поведение, направленное на прием воды.
- Центры регуляции деятельности желудочно-кишечного тракта и пищевого поведения: в латеральном гипоталамическом ядре – центр голода, в вентромедиальном – центр насыщения.
- В гипоталамусе есть центры белкового, углеводного и жирового обмена, центры регуляции сердечно-сосудистой системы, проницаемости сосудов и тканевых мембран, регуляции мочеотделения.
- Гипоталамус участвует в регуляции сна и бодрствования (задний гипоталамус активизирует бодрствование, передний – сон).
- Регуляция эмоционального поведения (раздражение заднего гипоталамуса вызывает активную агрессию, а передних отделов – пассивно-оборонительную реакцию, страх, ярость); центр полового поведения.
Особое место в функциях гипоталамуса занимает регуляция деятельности гипофиза. Благодаря гипоталамо-гипофизарным связям гипоталамус является высшим центром эндокринной регуляции.
Источник
Функционально в нем выделяют 2 отдела: таламус и гипоталамус. В таламусе происходит обработка почти всей информации, идущей от рецепторов к коре. Через него проходят сигналы от зрительных, слуховых, вкусовых, кожных, мышечных, висцеральных рецепторов, а также ядер ствола мозга, мозжечка, подкорковых. Сам он содержит около 120 ядер. Они делятся: на неспецифические и специфические. Неспецифические относятся к переднему отделу ретикулярной формации ствола мозга. Их аксоны нейронов поднимаются к коре и диффузно пронизывают все ее слои. К этим ядрам подходят нервные волокна от нижележащих отделов Р.Ф., гипоталамуса, лимбической системы, базальных ядер. При возбуждении неспецифических ядер в коре мозга развивается периодическая электрическая активность в виде веретен, что свидетельствует о переходе к сонному состоянию. Т.е. они обеспечивают определенный уровень функционального активности коры.
Специфические ядра делятся на переключающие или релейные и ассоциативные. Переключающие ядра состоят из нейронов, у которых мало дендритов и длинный аксон. С помощью них происходит переключение сигналов идущих от нижележащих отделов ЦНС на соответствующие соматосенсорные зоны коры, в которых находится представительство определенных рецепторов. Например в латеральных коленчатых телах переключаются зрительные сигналы на затылочные доли коры. В . переключающих ядрах выделяется наиболее важная информация. При нарушении функции этих ядер выключается восприятие соответствующих сигналов.
Ассоциативные нейроны имеют большее количество отростков и синапсов. Это позволяет им воспринимать различные по характеру сигналы. Они их получают эти сигналы от переключающих и осуществляют их первичный синтез. От них пути идут к ассоциативным зонам коры, в которых происходит высший синтез и формируются сложные ощущения.
Кроме того, ядра таламуса участвуют в формировании безусловных двигательных рефлексов сосания, жевания, глотания. В таламусе находится подкорковый центр болевой чувствительности, в котором формируется общее ощущение боли, не имеющее определенной локализации и окраски.
В гипоталамусе выделяют 32 пары ядер. Их несколько групп – преоптические, передние, средние, наружные и задние. Гипоталамус имеет многочисленные восходящие связи с лимбической системой, базальньши ядрами, таламусом, корой. Нисходящие пути от него идут к таламусу, ретикулярной формации, вегетативным центрам ствола и спинного мозга.
Гипоталамус является высшим подкорковым центром вегетативной регуляции. На висцеральные функции организма он влияет двумя путями. Во-первых через вегетативную нервную систему. Его передние ядра являются высшими парасимпатическими центрами. Поэтому при их возбуждении урежаются сердцебиения, снижается АД, понижается энергетический обмен, температура тела, суживаются зрачки и т.д. При возбуждении задних ядер возникает обратная картина, т.к. они являются высшими симпатическими центрами. Во-вторых, ГТ влияет на многие функции через гипофиз. Посредством нервных и сосудистых связей он образует с ним единую гипоталамо-гипофизарную систему. Такое взаимодействие связано с тем, что некоторым нейронам ГТ свойственно явление нейросекреции. Это способность продуцировать гормоноподобные вещества. В частности, в супраоптическом ядре вырабатываются нейрогормоны вазопрессин и окситоцин. По аксонам секретирующих нейронов они поступают в заднюю долю гипофиза, а оттуда выделяются в кровь. В медиальных ядрах синтезируются либерины и статины. По венозной гипоталамо-гипофизарной сети они транспортируются к передней доле гипофиза. Первые стимулируют синтез и выделение его гормонов, вторые тормозят. В свою очередь тропные гормоны влияют на функции других желез внутренней секреции.
Благодаря многочисленным связям, высокой чувствительности нейронов гипоталамуса к составу омывающей его крови, отсутствию в этом отделе гематоэнцефалического барьера, в нем находятся центры терморегуляции, регуляции водно-солевого обмена, обмена белков, жиров, углеводов и др. За счет них регулируется гомеостаз.
Гипоталамус участвует в формировании некоторых мотиваций и поведенческих реакций. Например, мотиваций и поведения голода, жажды. При раздражение вентромедиального ядра чувство голода и соответствующее поведение исчезают. При его разрушении наоборот наступает неутолимый голод. Т.е. здесь находятся центры голода и насыщения. При раздражении паравентрикулярного ядра развивается чувство жажды и питьевое поведение, а при разрушении жажда исчезает. В гипоталамусе расположены центры бодрствования и сна. В опытах с самораздражением (Олдс), когда в определенные ядра ГТ вживляются электроды, установлено, что здесь находятся центры двух базисных эмоций -удовольствия и неудовольствия. При раздражении некоторых ядер ГТ у человека возникает эйфория, повышается сексуальность.
ГТ принадлежит важная роль в развитии стресса, т.е. реакций напряжения на угрожающую ситуацию. При воздействии физиологических или психологических стрессоров (холод, недостаток кислорода, эмоциональном напряжении) кора посылает сигналы к симпатическим центрам ГТ, которые активируют симпатический отдел вегетативной нервной системы, выделение кортикотропинрелизинг гормона, а как следствие АКТГ. В результате происходит симпатическая активация внутренних органов, выделяются адреналин из мозгового слоя и кортикостероиды.
При патологии ГТ возникают расстройства терморегуляции (гипер- и гипотермия), аппетита (афагия-, гиперфагия), сна. Эндокринные нарушения, связанные с гипоталамусом, могут проявляться преждевременным половым созреванием, нарушениями менструального цикла, полового влечения, несахарным диабетом.
Соседние файлы в предмете [НЕСОРТИРОВАННОЕ]
- #
- #
- #
- #
- #
- #
- #
- #
- #
- #
- #
Источник