Центр мочеиспускания расположен в продолговатом

Высшие центры контроля мочеиспускания: цикл мочеиспускания, его нарушения
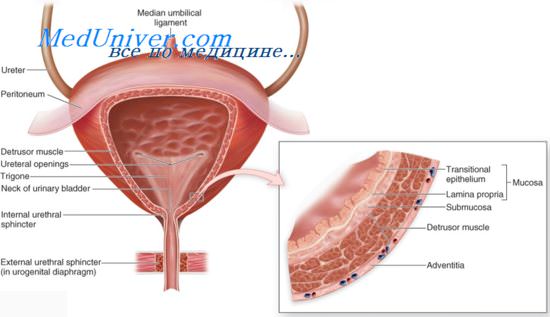
Нижние мочевыводящие пути разделены на два отдела—вместилище мочи (мочевой пузырь), а также собственно выводящие пути—уретру и наружный сфинктер уретры, действующих скоординировано для хранения и выведения мочи. На уровне спинного мозга парасимпатические нейроны крестцовых сегментов вызывают сокращение мочевого пузыря (детрузора). Симпатические нейроны поясничных сегментов угнетают парасимпатические влияния и позволяют мочевому пузырю наполниться.
Двигательные нейроны крестцовых сегментов, иннервирующие наружный сфинктер уретры, вносят вклад в функционирование обеих систем: при их возбуждении облегчается накопление мочи, при их торможении—облегчается ее выведение. Взаимодействие между этими системами происходит на различных уровнях ЦНС.
Мостовой центр контроля мочеиспускания (ядро Баррингтона) находится в парамедианной мостовой ретикулярной формации с каждой стороны, соединяясь по срединной линии. Отсюда отходят волокна крупноклеточных нейронов к отвечающим за мочеиспускание парасимпатическим нейронам в сегментах S2-S4 спинного мозга. Возбуждение центра контроля мочеиспускания приводит к акту мочеиспускания посредством повышения внутрипузырного давления (в результате сокращения гладких мышц стенки мочевого пузыря) и одновременного расслабления поперечно-полосатых мышц наружного уретрального сфинктера (вследствие возбуждения ГАМК-ергических вставочных нейронов ядра Онуфа, образующих синапсы с крестцовыми сегментами спинного мозга).
Аксоны этих двигательных нейронов проходят в составе половых нервов, иннервирующих наружный уретральный сфинктер. (В более латеральном отделе ретикулярной формации моста расположена L-зона, имеющаяся у млекопитающих, от которой волокна направляются к ядру Онуфа и вызывают сокращение наружного сфинктера уретры. Мостовой центр контроля мочеиспускания обозначают как М-зону, однако его взаимодействие с L-зоной вызывает сомнения.)
На более высоких уровнях к клеткам латерального отдела правого ОВСВ подходят восходящие волокна от задних рогов крестцовых сегментов, а от них—возбуждающие волокна к островку, который генерирует чувство наполненности мочевого пузыря и передает возбуждение на медиальные отделы лобной коры. Возбуждение латерального отдела ОВСВ осуществляет также правый гипоталамус. Часть спиноретикулярных волокон от крестцовых сегментов возбуждает L-зону. Другие волокна переключаются через таламус на клетки правой передней поясной коры (ППК), возбуждающиеся в ситуациях, требующих концентрации внимания.
Такое доминирование правых отделов может быть связано с эмоциональными аспектами мочеиспускания. При функциональном исследовании мозга было установлено, что дальнейшее заполнение жидкостью уже полного мочевого пузыря приводит к активации островка и дорсальной передней поясной коры/дополнительной моторной области. Когда мочевой пузырь не наполнен, заполнение жидкостью приводит к активации среднего мозга и парагиппокампальной области, осуществляющих бессознательный контроль афферентных сигналов от мочевого пузыря.
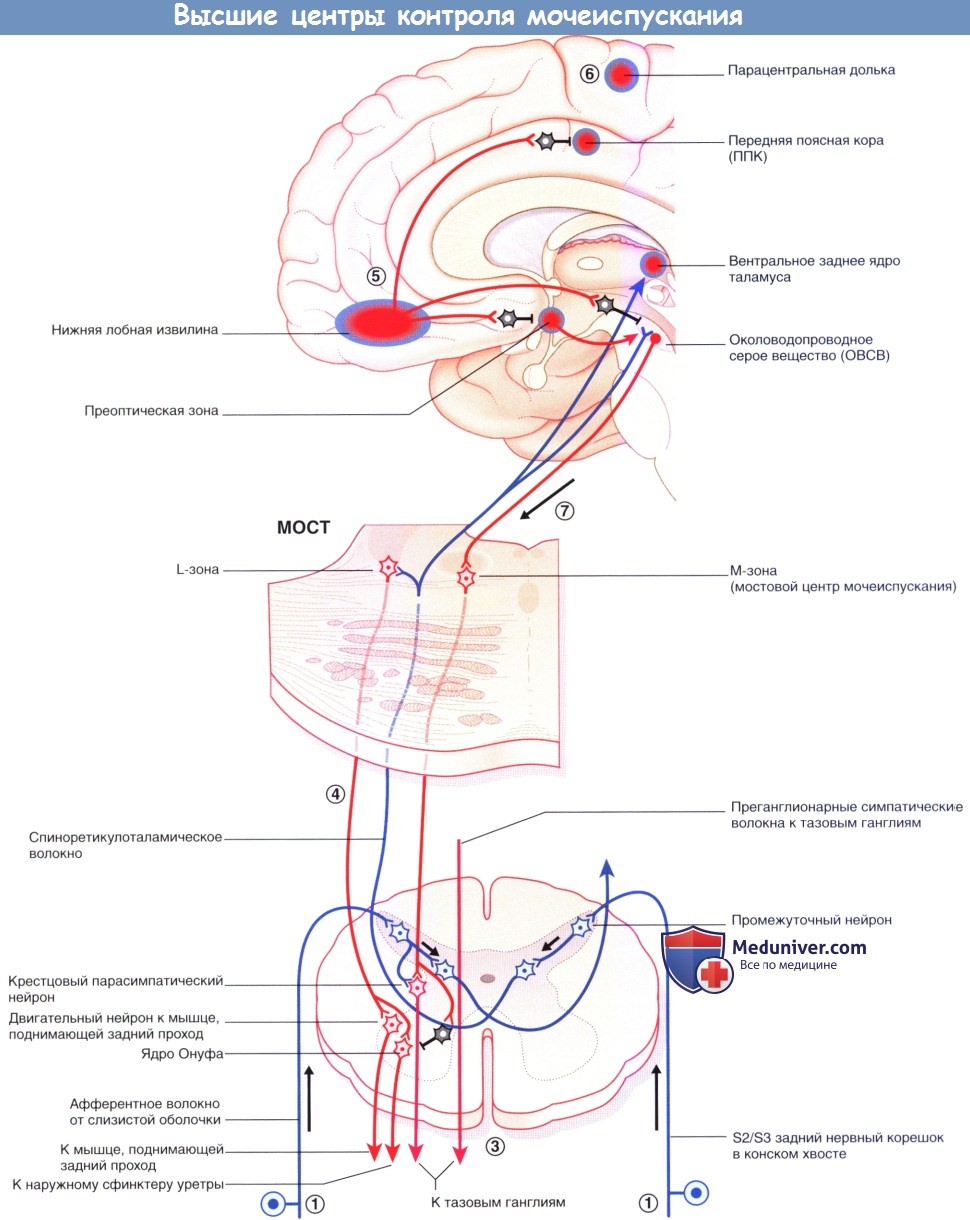
а) Цикл мочеиспускания:
1. Когда мочевой пузырь заполнен наполовину, восходящие волокна от рецепторов растяжения в детрузоре и слизистой оболочке в мочепузырном треугольнике передают эту информацию по спиноретикулярным волокнам через таламус к мосту, среднему мозгу и мозжечку.
2. От коры островка информация поступает в центр принятия решений в медиальной лобной коре, которая получает данные о степени наполнения мочевого пузыря.
3. Активация симпатической системы увеличивается постепенно, позволяя увеличить растяжимость мочевого пузыря (через β2-адренорецепторы). При взаимодействии с α2-адренорецепторами происходит торможение парасимпатических нейронов.
4. Спиноретикулярные волокна, образующие синапсы с L-зоной моста, возбуждают ядро Онуфа в крестцовых сегментах спинного мозга, повышая, таким образом, тонус наружного сфинктера уретры.
5. При полном заполнении возникает ощущение непреодолимого желания. При неподходящем времени или месте активируется отдел медиальной лобной извилины. Эта область тормозит ППК, уменьшая уровень ее активности путем возбуждения ее тормозных вставочных нейронов. Аналогичным образом через переключение в гипоталамусе и среднем мозге происходит торможение пре-оптической области и ОВСВ посредством возбуждения тормозных нейронов.
6. Последняя, но кратковременная мера —произвольное сокращение мышц тазовогодна. Команду на сокращение посылает лобная кора к промежностному отделу в медиальной области моторной коры парацентральной дольки.
7. Когда время и место позволяет, медиальная лобная извилина «освобождает троих своих пленников». Происходит расслабление тазового дна, гипоталамус активирует М-зону ОВСВ, одновременно с этим ингибируя L-зону путем возбуждения тормозных вставочных нейронов. Правостороннее доминирование центра контроля мочеиспускания согласуется с клиническими данными: у пациентов обоих полов, перенесших инсульт, недержание мочи чаще возникает при правосторонней локализации очага.
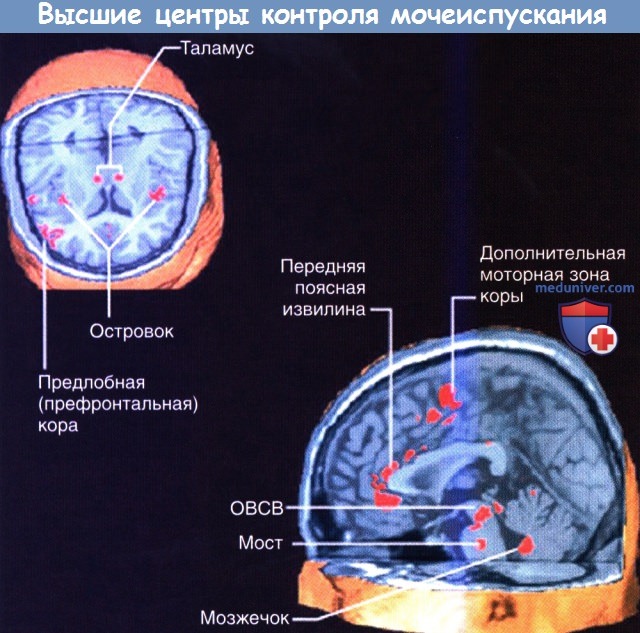
Показаны высокоактивные области при функциональной магнитно-резонансной томографии (фМРТ) в фазу наполнения мочевого пузыря в цикле мочеиспускания.
ОВСВ — околоводопроводное серое вещество. Мост — подразумевается мостовой центр контроля мочеиспускания.
б) Роль моноаминов. К двигательным и чувствительным ядрам спинного мозга, иннервирующим мочевой пузырь, подходит огромное количество серотонинергических нисходящих волокон от большого ядра шва (БЯШ) продолговатого мозга. Доказано, что растяжение мочевого пузыря приводит к стимуляции БЯШ (путем спиноретикулярной активации ОВСВ). Кратко излагая, контроль мочеиспускания на низшем уровне означает, что БЯШ создает тонус мочевого пузыря, направленный, прежде всего, на его наполнение.
Норадренергические нисходящие волокна из голубого пятна к передним рогам спинного мозга усиливают действие глутамата, локально вырабатываемого из клеток ядра Онуфа, увеличивая, тем самым, тонус наружного сфинктера в фазу наполнения.
в) Императивное недержание мочи. Императивное недержание мочи — неспособность взрослой женщины контролировать опорожнение мочевого пузыря до завершения фазы наполнения цикла мочеиспускания. Оно характеризуется неотложным позывом, быстро переходящим в неконтролируемое мочеиспускание, независимо от обстоятельств (отсюдатермин — «гиперактивный мочевой пузырь», или «гиперактивность детрузора»), Во многих случаях в анамнезе имеется указание на раздраженный мочевой пузырь в детском возрасте в форме частого дневного мочеиспускания и/или ночного энуреза (ночного недержания мочи).
При функциональном исследовании мозга (фМРТ) у здорового взрослого человека можно обнаружить повышение активности правой островковой области, которая, вероятно, отвечает за повышенную реактивность мочевого пузыря, и передней (отвечающей за эмоции) области поясной коры, обусловливающей возникновение неотложного позыва и «страх» неизбежного мочеиспукания.
G-белковые мускариновые рецепторы, активируемые постганглионарными волокнами тазовых ганглиев, в большом количестве присутствуют в детрузоре мочевого пузыря. Соответственно, препаратами выбора в данном случае служат блокаторы м-холинорецепторов. Однако появление антихолинергических побочных эффектов, таких как сухость во рту и запор, может потребовать отмены терапии.
В последние годы для лечения гиперактивности детрузора все чаще используют ботулотоксин. Известно, что он нарушает взаимодействие между холинергическими синаптическими везикулами и мышечными волокнами (как гладкими, так и поперечно-полосатыми), делая синаптическую передачу неэффективной. Гибкий цистоскоп проводят через уретру и выполняют многочисленные небольшие инъекции ботокса в стенку мочевого пузыря. Для получения долговременного результата процедуру повторяют 2 раза в год.
– Вернуться в оглавление раздела “Неврология.”
Редактор: Искандер Милевски. Дата публикации: 20.11.2018
Источник
Рефлекс мочеиспускания. Воздействие головного мозга на мочеиспусканиеНа рисунке показано, что на относительно постоянном фоне давления по мере наполнения пузыря возникает множество быстрых колебаний в виде мочеиспускательных сокращений (изображены на графике прерывистыми линиями). Эти сокращения возникают рефлекторно, стимулом для рефлекса служит активация рецепторов растяжения стенки пузыря, особенно отдела задней уретры, который начинает заполняться при высоком внутрипузырном давлении. Чувствительные импульсы от рецепторов растяжения распространяются в крестцовые сегменты спинного мозга по тазовым нервам, возвращаясь оттуда к пузырю по парасимпатическим порциям тех же нервов. Мочеиспускательные сокращения при частичном заполнении пузыря в течение нескольких секунд обычно самопроизвольно приводят к расслаблению детрузора, и давление возвращается к исходному уровню. Поскольку пузырь продолжает наполняться, рефлекс мочеиспускания возникает все чаще, а сила сокращений детрузора увеличивается. Возникнув однажды, рефлекс мочеиспускания становится самоподдерживаемым, т.е. первичное сокращение пузыря активирует рецепторы растяжения, поток чувствительной импульсации от пузыря и задней уретры нарастает, усиливая рефлекторное сокращение стенки. Цикл повторяется вновь, пока мочевой пузырь не достигнет высокой степени сокращения. Затем через несколько секунд (иногда проходит более 1 мин) самоподдерживаемый рефлекс мочеиспускания угасает, позволяя пузырю расслабиться. Итак, рефлекс мочеиспускания является завершенным циклом реакций, состоящим из следующих периодов: (1) постепенно и быстро возрастающего давления; (2) стойкого поддержания давления; (3) возврата давления к банальному тонусу мочевого пузыря. Рефлекс мочеиспускания, возникнув и не завершившись изгнанием мочи, затормаживается, пребывая в таком состоянии от нескольких минут до 1 ч и более, вплоть до наступления следующего цикла. Поскольку мочевой пузырь наполняется все сильнее, частота и выраженность рефлекса мочеиспускания возрастают все больше. Став достаточно сильным, рефлекс мочеиспускания запускает другую рефлекторную реакцию, которая реализуется с помощью тормозного влияния срамных нервов на наружный сфинктер уретры. Если торможение будет преобладать над произвольными сигналами ЦНС, увеличивающими тонус наружного сфинктера, произойдет мочеиспускание. В противном случае мочевой пузырь будет наполняться до тех пор, пока рефлекс мочеиспускания не станет более выраженным.
Воздействие головного мозга на мочеиспусканиеРефлекс мочеиспускания относится к вегетативным рефлексам спинного мозга, однако его можно подавить или усилить с помощью высших отделов головного мозга, которые включают: (1) столовые центры, вызывающие стойкое возбуждение и торможение, расположенные преимущественно в мосте (2) несколько центров, расположенных в коре и в основном выполняющих тормозную функцию, иногда способных оказывать и возбуждающее действие. Рефлекс мочеиспускания является основой одноименного процесса, окончательная роль в его регуляции, однако, принадлежит высшим отделам ЦНС. 1. Центры, расположенные в головном мозге, способствуют постоянному слабому торможению рефлекса мочеиспускания. Торможение прекращается лишь при осознанном желании помочиться. 2. Высшие центры способны предотвратить мочеиспускание даже при начавшемся опорожнении пузыря посредством тонического сокращения наружного сфинктера, которое продолжается вплоть до подходящего момента, когда мочеиспускание может быть вновь продолжено. 3. Корковые влияния могут содействовать активации центра мочеиспускания, расположенного в крестцовом отделе, и одновременному с ним торможению сокращения наружного сфинктера уретры, что может привести к мочеиспусканию наполненного пузыря. Произвольное мочеиспускание обычно вызвано следующими причинами: произвольным сокращением брюшной стенки, которое способствует повышению давления внутри пузыря, попаданию мочи в область шейки и задней уретры, растяжению стенок, возбуждению механорецепторов, активации рефлекса мочеиспускания и одновременному торможению наружного сфинктера уретры. При мочеиспускании, как правило, пузырь опорожняется целиком, за редким исключением в нем остается более 5—10 мл мочи. – Также рекомендуем “Атония мочевого пузыря. Непроизвольный мочевой пузырь” Оглавление темы “Физиология почек и их функция”: |
Источник
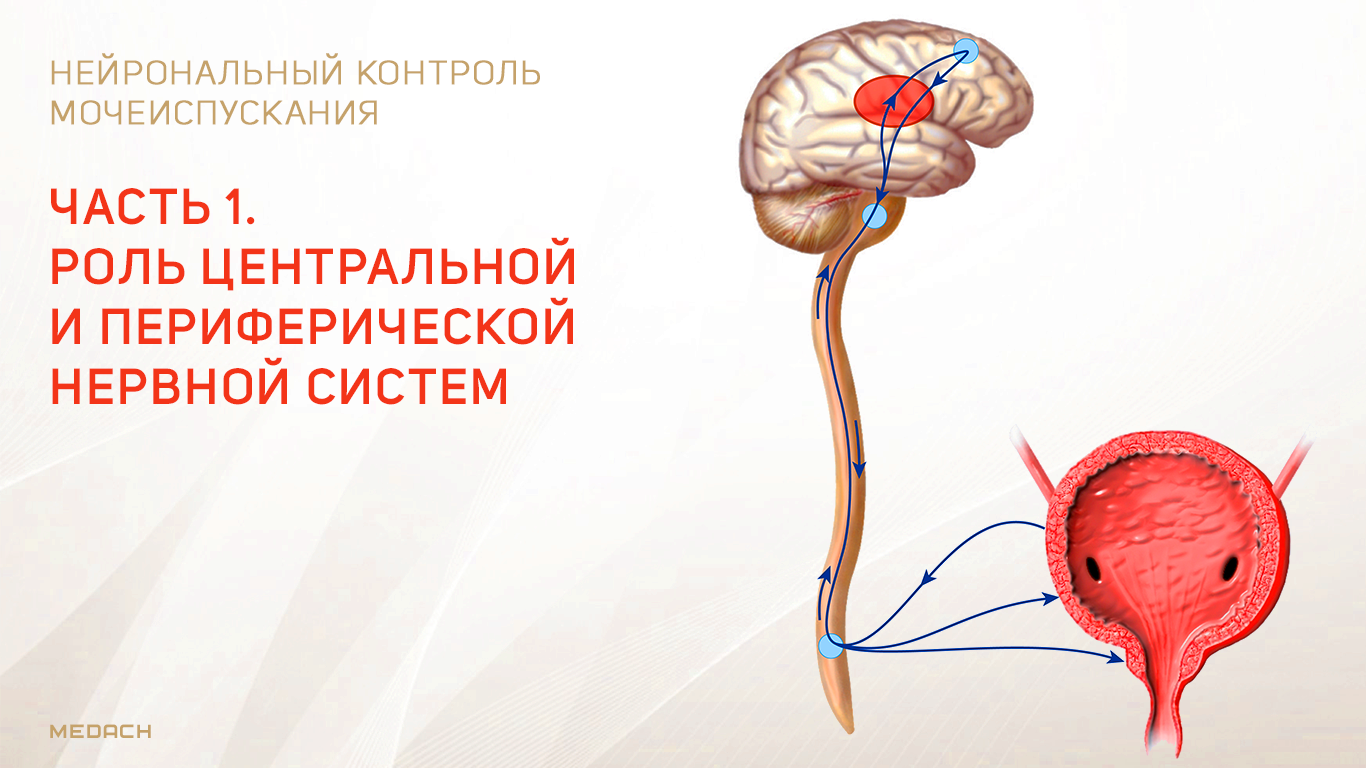
Хранение мочи и периодическое опорожнение мочевого пузыря (МП) зависят от координированной работы гладких и поперечнополосатых мышц, условно разделенных на два структурно функциональных отдела: резервуара (мочевой пузырь) и выпускной системы, состоящей из шейки МП, уретры и наружного сфинктера мочеиспускательного канала. За координированную работу вышеперечисленных структур отвечают сложные нейронные системы управления, расположенные в периферических ганглиях, спинном и головном мозге.
Обеспечение сознательного контроля мочеиспускания требует сложных взаимодействий между вегетативным (симпатика и парасимпатика) и соматическим отделами нервной системы.
Симпатические нервные волокна берут свое начало из боковых рогов грудопоясничного отдела (спинномозговой центр Якобсона) и ганглиях нижнего брыжеечного сплетения и достигают МП в составе подчревного нерва. Симпатические постганглионарные нервы выделяют норадреналин, который активирует β-адренергические рецепторы, ингибирующие сокращение детрузорной мышцы, и α-адренергические рецепторы, возбуждающие мускулатуру уретры и шейки МП. Симпатическое воздействие приводит к расслаблению мышцы, изгоняющей мочу, сокращению мускулатуры уретры и шейки МП и ингибированию интрамуральных ганглиев МП.
Центральный отдел парасимпатической иннервации МП располагается в промежуточных ядрах крестцовых сегментов. Холинергические преганглионарные волокна из промежуточных ядер посылают свои аксоны через тазовые нервы к ганглиозным клеткам тазового сплетения и интрамуральным нейронам в стенке мочевого пузыря. Ганглиозные клетки в свою очередь возбуждают детрузор мочевого пузыря, что приводит к его сокращению с последующим опорожнением. Основными медиаторами парасимпатической нервной системы тут являются ацетилхолин и другие нехолинергические медиаторы. Ацетилхолин действует опосредованно, возбуждая M3-холинорецепторы клеток детрузора. Парасимпатические нервные окончания в нервно-мышечных синапсах и в парасимпатических ганглиях тоже имеют холинорецепторы, возбуждение этих рецепторов на нервных окончаниях может усиливать (через рецепторы М1) или подавлять (через рецепторы М4) высвобождение медиаторов в зависимости от интенсивности нейронного возбуждения. Основной нехолинергический медиатор это АТФ, который активирует внутриклеточную пуринергическую систему через возбуждение P2X рецепторов и тоже способствует сокращению детрузора. Парасимпатические волокна вызывают расслабление ГМК уретры путем высвобождения оксида азота (NO).
Аксоны соматических двигательных нейронов передних рогов крестцовых сегментов S2–S4 (ядро Онуфа) проходят в срамном нерве и иннервируют поперечно-полосатые мышцы наружного уретрального сфинктера. Нейроны более медиально расположенного моторного ядра на том же уровне позвоночника иннервируют мускулатуру тазового дна.
Афферентные пути НМП состоят из цепочек чувствительных нейронов. Первые нейроны, располагающиеся в спинальных ганглиях на уровне S2–S4 и T11–L2, реагируют на пассивное растяжение и активное сокращение мышц МП и передают эту информацию на нейроны второго и третьего порядка. Эти нейроны обеспечивают координированную работу спинальных рефлексов и восходят к вышележащим структурам головного мозга, контролирующих фазы накопления и опорожнения МП. Наиболее важные афферентные волокна от мочевого пузыря идут в составе тазового нерва, в то время как чувствительность от шейки МП и уретры передается по срамным и подчревным нервам. Афферентные волокна этих нервов состоят из миелинизированных (Аδ) и немиелинизированных (С) аксонов. Aδ-волокна передают информацию о наполнении мочевого пузыря. С-волокна нечувствительны к изменению объема мочевого пузыря в физиологических условиях, поэтому они называются «тихими». Они реагируют главным образом на патологические стимулы, такие как химическое раздражение или охлаждение.
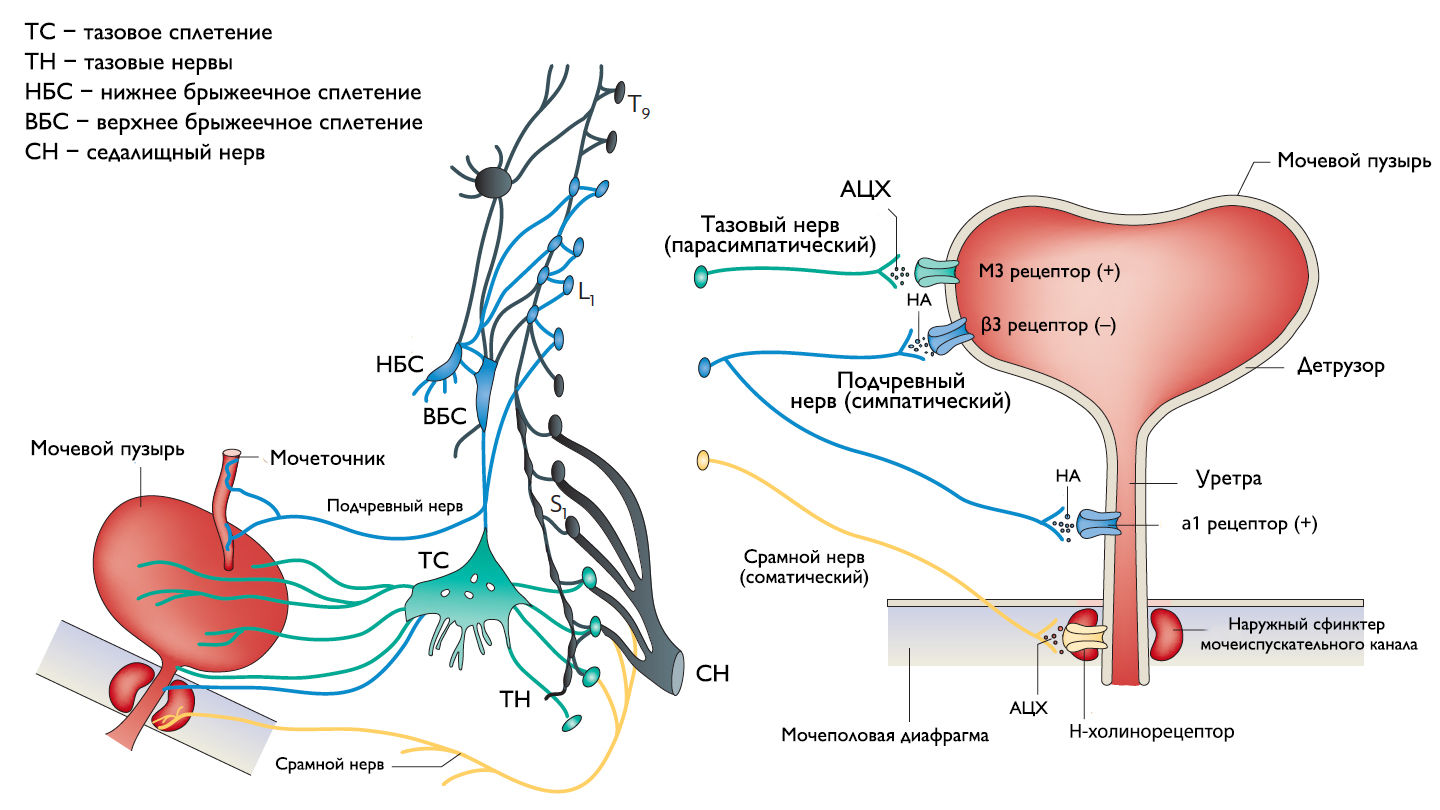
Рисунок 1 | Иннервация нижних мочевыводящих путей.
Специфический и неспецифический восходящий супраспинальный сенсорный путь
Одни спинномозговые промежуточные нейроны посылают восходящие волокна к определенным областям моста и среднего мозга, участвующим в мочеиспускании. Другие промежуточные нейроны передают информацию из нижних мочевыводящих путей в структуры переднего мозга, включая таламус и гипоталамус. Спиноталамический и спиногипоталамический тракты хотя и не играют главную роль в мочеиспускании, но могут включаться в сознательный контроль полноты мочевого пузыря. Чувствительные зоны коры ГМ через спиноталамический тракт информируются о состоянии наполнения МП.
Мостовой центр мочеиспускания (МЦМ) и его нисходящие спинномозговые двигательные пути
Впервые центр управления мочеиспусканием был открыт в дорсальной части моста Баррингтоном в 1925 году и с тех пор называется мостовым центром мочеиспускания или ядром Баррингтона. МЦМ располагается в области покрышки моста. Нейроны МЦМ имеют нисходящие возбуждающие синаптические контакты с клетками парасимпатических преганглионарных мотонейронов, иннервирующих постганглионарные клетки мочевого пузыря. Электрическая и химическая стимуляция МЦМ у крыс и кошек инициирует сокращение мочевого пузыря и расслабляет сфинктер уретры имитируя нормальное мочеиспускание. Цикл рефлекса мочеиспускания состоит из трех фаз, контролируемых различными центральными механизмами: фаза реализации безопасной среды — для начала мочеиспускания человеку необходимо осознание, что окружающие обстановка комфортна; фаза релаксации наружного уретрального сфинктера; и фаза сокращения мышцы, выталкивающей мочу. Процесс нормального мочеиспускания невозможен без какой-либо из этих фаз. МЦМ является командным центром мочеиспускания, который контролирует последовательное переключение фазы расслабления наружного уретрального сфинктера на фазу сокращения детрузорной мышцы.
Мостовой центр удержания мочи (МЦУ) и его нисходящие спинномозговые двигательные пути
Его роль заключается в расслаблении детрузора и сокращении наружного уретрального сфинктера. МЦУ располагается вентролатеральней МЦМ. Синапсы волокон МЦУ возбуждают ядро Онуфа в крестцовых сегментах спинного мозга, повышая таким образом тонус наружного сфинктера уретры. Стимуляция области МЦУ останавливает мочеиспускание, возбуждает мышцы тазового дна и сокращает уретральный сфинктер. Наоборот, двусторонние поражения МЦУ вызывают недержание мочи, чрезмерную детрузорную активность, невозможность хранения мочи и снижение тонуса уретрального сфинктера. На сегодняшний день нет анатомических доказательств связи между МЦУ и МЦМ, и было высказано предположение, что эти центры функционально независимы.
Роль кортикальных областей
Наиболее частые симптомы поражения кортикальных областей ГМ это поллакиурия и ургентное недержание мочи. Поэтому Andrew и Nathan выдвинули гипотезу, что отсоединение лобной или передней поясной извилины от гипоталамуса приводит к непроизвольному началу мочеиспускания [3]. Действительно, префронтальная кора головного мозга человека и передняя поясная извилина активируются во время мочеиспускания [4].
Мозжечок и базальные ганглии
Существует ряд исследований о том, что мозжечок и базальные ганглии оказывают в основном ингибирующее действие на мочевой пузырь. Мозжечковая патология приводит к увеличению частоты мочеиспускания и ургентному недержанию мочи. Симптомы гиперактивного мочевого пузыря также встречаются при болезни Паркинсона. Поскольку нет прямых связей этих областей с МЦМ, ингибирующее влияние, вероятно, косвенное через структуры переднего и среднего мозга.
Рисунок 2 | Предположительное схематическое изображение связей между различными структурами переднего мозга и ствола мозга, которые участвуют в контроле мочеиспускания.
- Liao L., Madersbacher H. (ed.). Neurourology: Theory and Practice. — Springer, 2019.
- Clare J. Fowler et al. The neural control of micturition. — Nature Reviews | Neuroscience, volume 9. — June 2008.
- Andrew J, Nathan PW. Lesions of the anterior frontal lobes and disturbances of micturition and defecation. Brain. 1964;87:233–62.
- Griffiths, Derek J. “Use of functional imaging to monitor central control of voiding in humans.” Urinary Tract. Springer, Berlin, Heidelberg, 2011. 81-97.
Нашли опечатку? Выделите фрагмент и нажмите Ctrl+Enter.
Источник