Амитоз в клетках мочевого пузыря лягушки

Препарат представляет собой гистологический срез мочевого пузыря мыши, окрашенный по Нонизеду и Уиндлю (Микрофото 5). (рис. 22, 23)
При малом увеличении видны: ткань построенная из клеток с хорошо различимыми оболочками и ядрами различной величины и формы (от округлой до гантелеобразной) ядра эпителиальных клеток слизистой оболочки мочевого пузыря. Слабо оксифильная цитоплазма этих клеток почти неразличима, одинаково с опущенным конденсором и прикрытой диафрагмой можно видеть клетки различной величины. Передвигая препарат, отыскиваем делящиеся клетки, где ядро сильно вытянуто в длину, в средней его части образуется перетяжка, которая истончается. Видны места, где клетка становится двуядерной, места, где есть разделение двуядерной клетки на две одноядерные (процесс цитотомии или плазмотомии). Встречаются клетки многоядерные в результате задержания цитотомии или при отсутствии ее. В крупных, неправильной округлой формы клетках содержится одно, два, три и более ядер. Среди неделящихся ядер можно видеть также ядра, находящиеся на различных стадиях амитотического деления. Эти ядра надо изучить при большом увеличении.
Рис. 23. Амитоз эпителиальных клеток. Отпечаток поверхности слизистой оболочки мочевого пузыря мыши. 1 – ядро, 2 – перетяжка ядра, 3- двуядерная клетка, 4 – цитомия, 5 – многоядерные клетки.
Во время прямого деления ядро остается в интерфазном состоянии и клетка не перестает функционировать. О начале амитоза свидетельствует изменение формы ядра. Основа вытягивается в длину и разрывается. Клетка становится двуядерной. В дальнейшем может произойти цитотомия. Нередко цитотомия задерживается или вообще не наступает, в результате чего возникают многоядерные клетки.
Иногда видно деление ядрышка, которое удлиняется и перешнуровывается. Прямое деление клеток эпителия мочевого пузыря является дегенеративной формой, так как предшествует отмиранию этих клеток.
Обозначения: 1 – делящиеся клетки, 2 – двуядерные клетки, 3 – перетяжка ядра и цитоплазмы, 4 –многоядерные клетки.
Задания
1. Изучить деления клеток.
2. Изучить амитоз.
Контрольные вопросы:
- Какова общая характеристика амитоза.
- Биологическое значение амитоза.
ЗАНЯТИЕ 3
Тема 3. ПЛАЗМАТИЧЕСКАЯ МЕМБРАНА (ПЛАЗМАЛЕММА)
Содержание. Модели биологических мембран. Жидкостно-мозаичная модель. Особенности химического состава и его взаимосвязь со свойствами и функциями мембран. Клеточные стенки (оболочки). Плазмодесмы и десмосомы. Синцитий, симпласт, плазмодий.
Средства наглядности. Таблицы с изображением жидкостно-мозаичной модели мембраны
Оборудование и материалы. Препараты: кариотип традесканции. Таблицы с изображением кариотипов. Семена культурных злаков, луковицы, черенки ив
Сосуды емкостью 250 и 1000 мл, чашки Петри, фильтровальная бумага, проволока, вода дистиллированная.
Задания для аудиторной работы
1. Законспектируйте теоретическую часть занятия.
2. Ответьте на контрольные вопросы.
3. Заполните таблицу «Химический состав клеточных оболочек».
Задания для внеаудиторной работы
Продолжите изучение основных классов органических соединений клетки. Обратите внимание на структуру и функции углеводов, липидов и белков.
Теоретическая часть
Плазматическая мембрана (цитоплазматическая мембрана), или плазмалемма.
Существование плазмалеммы предсказал Франц Мейен (1830), который считал, что клетка – это пространство, отграниченное от внешней среды вполне замкнутой мембраной.
Овертон (начало ХХ века) установил, что плазматическая мембрана эритроцитов содержит большое количество липидов. Гортер и Грендел (1925) доказали, что мембрана состоит из двойного слоя липидов (липидный бислой). Доусон и Даниелли (1935) предложили модель бутерброда: мембрана состоит из липидного бислоя, заключенного между двумя слоями белка. Робертсон (1959) на основе анализа данных электронной микроскопии установил, что все мембраны построены по единому плану: липидный бислой + белковые молекулы. Сингер и Николсон (1972) разработали жидкостно-мозаичную модель мембраны, которая является в настоящее время общепринятой.
Согласно жидкостно-мозаичной, или жидкокристаллической модели, основу мембран составляет фосфолипидный бислой. Гидрофильные глицерофосфатные части молекул фосфолипидов находятся на внешних поверхностях бислоя. Гидрофобные углеводородные части молекул фосфолипидов направлены вовнутрь бислоя. Структура бислоя поддерживается за счет поверхностного натяжения; связи между молекулами фосфолипидов называются гидрофобными. Отдельные блоки бислоя способны перемещаться относительно друг друга во всех направлениях.
Кроме фосфолипидов в состав мембран входят гликолипиды и стероиды (например, холестерин). Конкретный липидный состав мембран зависит от таксономической принадлежности организмов, от тканевой принадлежности клеток и от их физиологического состояния, а также от условий обитания организмов.
Белки мембран представлены простыми белками, гликопротеинами, липопротеинами, металлопротеинами и другими сложными белками. Выделяют три основных типа белков: периферические (гидрофильные, расположены на поверхности мембран), интегральные (гидрофобные, расположены в толще бислоя) и политопические (со смешанными свойствами, пронизывают мембрану насквозь). Белковые молекулы образуют непостоянные соединения между собой и небелковыми группами. В ходе химических взаимодействий конформация белков и их свойства существенно изменяются.
Углеводы в составе мембран обычно представлены гликопротеинами и гликолипидами. Основная часть углеводов плазмалеммы расположена на ее внешней стороне и образует гликокаликс.
Такая структура мембраны обеспечивает ее основное свойство: избирательную проницаемость. Функции плазмалеммы: барьерная, транспортная, энерготрансформирующая, информационно-сигнальная.
С внешней стороны мембраны часто формируются надмембранные структуры – клеточные оболочки, или клеточные стенки. Основные функции клеточных оболочек (стенок): механическая и защитная.
Основные типы клеточных оболочек:
1. У большинства низших эукариот (у водорослей, у низших грибов и грибоподобных организмов) клеточная стенка состоит из целлюлозы и гемицеллюлоз.
2. У высших грибов (Eumycota) клеточная стенка содержит хитин (реже – хитозан) и полимеры глюкозы – глюканы. У дрожжеподобных грибов хитина почти нет.
3. У высших растений первичная клеточная стенка состоит из целлюлозы. Вторичные стенки содержат суберин или лигнин. Смежные клетки разделены срединными пластинками из пектинов. У многих низших и высших растений в состав стенок входят минеральные вещества: кремнезем, известь и др.
4. У прокариот клеточная оболочка многослойная. Внутренний слой (собственно клеточная стенка) построен на основе муреина. Внешние слои имеют разнообразный химический состав. У многих видов имеется слизистая капсула из полисахаридов.
Источник
АМИТОЗ (amitosis; греческий отрицательная приставка a-, mitos — нить + -ōsis) прямое деление ядра — деление клеточного ядра на две или несколько частей без образования хромосом и ахроматинового веретена; при амитозе ядерная мембрана и ядрышко сохраняются и ядро продолжает активно функционировать.
Прямое деление ядра впервые описано Ремаком (R. Bemak, 1841); термин «амитоз» предложен Флеммингом (W. Flemming, 1882).

Рис. 1. Амитоз с последующей цитотомией в клетках новорожденных мышей: 1—6 — последовательные стадии деления клетки
Рис. 2. Амитоз с образованием двуядерной клетки
Обычно амитоз начинается с деления ядрышка, затем делится ядро. Деление его может протекать по-разному: либо в ядре появляется перегородка — так называемая ядерная пластинка, либо оно постепенно перешнуровывается, образуя два или несколько дочерних ядер. При помощи цитофотометрических методов исследования установлено, что примерно в 50% случаев амитоза ДНК равномерно распределяется между дочерними ядрами. В других случаях деление заканчивается появлением двух неравных ядер (мероамитоз) или множества мелких неодинаковых ядер (фрагментация и почкование). Вслед за делением ядра происходит деление цитоплазмы (цитотомия) с образованием дочерних клеток (рис. 1); если цитоплазма не делится, возникает одна дву- или многоядерная клетка (рис. 2).
Амитоз характерен для ряда высокодифференцированных и специализированных тканей (нейроны вегетативных ганглиев, хрящевые, железистые клетки, лейкоциты крови, клетки эндотелия кровеносных сосудов и другое), а также для клеток злокачественных опухолей.
Беншшгхофф (A. Benninghoff, 1922), исходя из функционального назначения, предложил различать три типа амитоза: генеративный, реактивный и дегенеративный.
Генеративный амитоз — это полноценное деление ядер, после которого становится возможным митоз (см.). Генеративный амитоз наблюдается у некоторых простейших, в полиплоидных ядрах (см. Хромосомный набор); при этом происходит более или менее упорядоченное перераспределение всего наследственного аппарата (например, деление макронуклеуса у инфузорий).
Сходная картина наблюдается при делении некоторых специализированных клеток (печени, эпидермиса, трофобласта и другое), где амитоз предшествует эндомитоз — внутриядерное удвоение набора хромосом (см. Мейоз); образующиеся в результате эндомитоз а полиплоидные ядра подвергаются затем амитозу.
Реактивный амитоз обусловлен влиянием на клетку различных повреждающих факторов — облучения, химических препаратов, температуры и другое. Он может быть вызван нарушениями обменных процессов в клетке (при голодании, денервации ткани и другое). Этот тип амитотического деления ядра, как правило, не завершается цитотомией и приводит к появлению многоядерных клеток. Многие исследователи склонны рассматривать реактивный амитоз как внутриклеточную компенсаторную реакцию, обеспечивающую интенсификацию метаболизма клетки.
Дегенеративный амитоз — деление ядра, связанное с процессами деградации или необратимой дифференциации клетки. При этой форме амитоза происходит фрагментация, или почкование, ядер, не связанная с синтезом ДНК, что в ряде случаев является признаком начинающегося некробиоза тканей.
Вопрос о биологическом значении амитоза окончательно не решен. Однако не возникает сомнений в том, что амитоз — это вторичное явление по сравнению с митозом.
См. также Деление клетки, Клетка.
Библиография: Клишов А. А. Гистогенез, регенерация и опухолевый рост скелетно-мышечной ткани, с. 19, Л., 1971; Кнорре А. Г. Эмбриональный гистогенез, с. 22, Л., 1971; Михайлов В. П. Введение в цитологию, с. 163, Л., 1968; Руководство по цитологии, под ред. А. С. Трошина, т. 2, с. 269, М. — Л., 1966; Bucher О. Die Amitose der tierischen und menschlichen Zelle, Protoplasmalogia, Handb. Protoplasmaforsch., hrsg. v. L. V. Heilbrunn u. F. Weber, Bd 6, Wien, 1959, Bibliogr.
Ю. Е. Ершикова.
Источник: Большая Медицинская Энциклопедия (БМЭ), под редакцией Петровского Б.В., 3-е издание
Рекомендуемые статьи
Источник
Амитоз — описание процесса
Амитоз — от греческого слова «митоз»(нить) и «а»(частица отрицания), то есть «не митоз». Прямое деление ядра клетки на две части. При этом не образуется веретено деления и хромосомы распределяются неравномерно.
На схеме можно посмотреть как выглядит данное деление:

Источник: na5.club
История открытия
Изучение клетки стало продвигаться вперед с появлением усовершенствованных микроскопов в 1830-х годах. Впервые амитоз как явление был описан немецким и польским ученым Робертом Ремаком (Robert Remak) в 1841 году на примере тканей животных. До этого другие исследователи уже рассмотрели деление клетки, но только Ремак первый догадался, что оно связано с делением ядра. Он считал, что клетки образуются только при помощи деления, а ядро находится в каждой из них.
Позже, в 1855 году Ремак и немецкий ученый Рудольф Вирхов (Rudolf Ludwig Karl Virchow) одновременно пришли к окончательному выводу, что новые клетки появляются лишь при делении уже существующих. Это дало развитие клеточной теории, выдвинутой Шлейденом и Шванном.
Термин «амитоз» придумал немецкий биолог Вальтер Флеминг (Walther Flemming) в 1882 году.
Основные черты процесса
Это явление бывает у полиплоидных, отживающих или клеток с патологией и ведет к образованию многоядерных клеток. В последнее время наука отрицает, что амитоз — нормальный процесс репродукции.
Более экономичный и менее сложный, чем митоз и мейоз, ведь не нужно ни удваивать ДНК, ни создавать веретено деления. Спирализация хромосом отсутствует.
По сравнению с митозом происходит намного быстрее, так как отсутствуют многие ядерные процессы.
Когда наблюдается амитоз
Он характерен для одноклеточных организмов, например инфузорий, а еще для высокоспециализированных клеток с ослабленной физиологической активностью и с разными дегенеративными процессами. Для обреченных на гибель либо при различных патологических процессах, таких как злокачественный рост опухолей, в том числе у человека и животных.
Для процесса нормальной репродукции такой вид деления как амитоз, по мнению ученых, не подходит, потому что при нем не может быть адекватного сохранения и дублирования генетического материала.
Мы можем наблюдать амитоз:
- при увеличении корневого чехлика;
- в эпителии;
- при росте лука;
- в рыхлой соединительной ткани;
- в хрящевой ткани;
- в мускулатуре;
- в зародышевых оболочках;
- при увеличении тканей водорослей;
- в эндосперме.
Характеристики амитоза
- не перестраивает клетку;
- отсутствует веретено деления;
- нет спирализации хроматина;
- не выявляются хромосомы;
- нет удвоения ДНК;
- неравномерное распределение генетического материала.
Особенностью амитоза является то, что клетка сохраняет свою функциональную активность, и ядро находится в интерфазном состоянии. Сохраняются также ядерная мембрана и ядрышко, образующиеся при делении клетки разного размера.
Встречается амитоз довольно редко, но ученые разделили его на несколько типов.
- Генеративный — ядра делятся полноценно и в дальнейшем возможен мейоз.
- Реактивный — деления не происходит, образуются многоядерные клетки. Это связано с химическим воздействием, радиоактивным облучением, критической температурой окружающей среды и другими повреждающими факторами.
- Дегенеративный — обусловлен деградацией или необратимой дифференциацией клетки. Ядро просто распадается на фрагменты.
Как протекает процесс
При амитозе мы наблюдем сужение цитоплазмы. Ядро визуально растягивается, а потом становится похоже на гантель. Далее оно разрывается на части и образуются два самостоятельных ядра. Цитоплазма сжимается и делит старую клетку на новые, причем они могут быть разного размера.
Значение амитоза
Этот способ деления клетки позволяет быстро восстановить поврежденные ткани у растений и животных, быстро размножиться одноклеточным организмам. Окончательного мнения ученых по поводу биологического значения амитоза пока нет.
Насколько полезной была для вас статья?
Рейтинг: 5.00 (Голосов: 4)
Выделите текст и нажмите одновременно клавиши «Ctrl» и «Enter»
Текст с ошибкой:
Расскажите, что не так
Источник
Общие замечания
Общий вес мочеполовой системы (apparatus urogenitalis) травяной лягушки составляет 1,8% веса всего тела. Однако в течение года половые органы испытывают столь значительные колебания в своем объеме и весе, что правильное представление о соотношении их с другими органами можно получить, только вскрывая лягушек в разное время года. Непосредственно перед размножением, в частности, полость тела самки бывает буквально до предела переполнена яйцами. После размножения вес половых органов резко падает. Внешним выражением эндокринного действия половой системы лягушек являются, как известно, вторично-половые признаки. У лягушек их сравнительно немного. В первую очередь это брачные мозоли самца и его голосовые мешки (см. главу I и IV). Интересны брачные бугорки самок (см. главу IV).
Органы выделения
Продукты обмена веществ удаляются из организма лягушки несколькими путями. Газы уходят через легкие и кожу. Не исключено, что продукт сгорания белков — мочевина также в некоторой степени может быть удаляема через кожу. Однако главный путь для ее удаления— мочевые органы (organa uropoetica). В то время как у высших позвоночных главным местом образования мочевины служит печень, у лягушки дело обстоит иначе. «При экспериментальном удалении печени у лягушки содержание в крови мочевины не отличается от нормы. Очевидно, у лягушек мочевина может образоваться в других органах. Лягушки выделяют при обычной температуре относительно больше мочи, чем человек: в день человек выделяет около одной пятидесятой своего веса, а лягушка около трети. Зимой деятельность почек почти прекращается. Напротив, возрастание температуры на 10° увеличивает их работу в 2-3 раза. Удельный вес мочи лягушек 1, 0015. Моча по своему характеру гипотоническая. В ней содержится (в процентах по отношению ко всему азоту мочи): аммиака 3, 2—15, 0%, мочевины 82—84% п мочевой кислоты до 0, 4%.
Органы выделения состоят из почек, их выносящих протоков и мочевого пузыря. Строение почек (rеnеs) у обоих полов одинаковое. Это два удлиненных (ширина содержится в длине 3—4 раза) плоских тела, располагающихся справа и слева по бокам позвоночника. Передние концы почек приходятся на границе между VII и VIII позвонками. Задняя граница приходится примерно на уровне середины хвостца. Нередко встречается асимметричное положение почек. У взрослых зеленых лягушек длина почки составляет 18— 25% длины тела. Наружный (боковой) край почки слегка изогнут и своей выпуклостью выдается в сторону, а внутренний (срединный) — слегка фестончатый или зазубренный. Поверхность почки, обращенная к спине, гладкая и ровная, а противоположная — неровная. Часть последней занята надпочечником. Большая часть брюшной поверхности почки покрыта брюшиной. Та часть срединного края, через которую в почку входят и выходят сосуды (артерии и вены), называется воротами почки (hiIusrenis). Со сравнительно-анатомической точки зрения почку взрослой лягушки надлежит считать первичной, или туловищной, почкой (mеsоnерhrоs). На брюшной стороне почки можно видеть, иногда даже под лупой, 120—250 неправильно разбросанных отверстий ресничных воронок, или нефростомов. Последние у взрослой лягушки сообщаются с выносящими почечными венами и служат для проведения лимфы из полости тела в венозную систему. Изучение внутреннего строения почки показывает, что она слагается из большого числа (до 5000; в среднем 2 000) мочевых трубочек, или канальцев, связанных соединительной тканью. Каждая такая трубочка (нефрон) состоит из ряда отделов: почечного, или Мальпигиева, тельца (corpusculum renis — с. Malpighii ), слагающегося из клубочка с Баумановой капсулой, от которого отходит первый отдел, или «шейка». Внутри последней наблюдаются мерцательные реснички. Потом. следует второй отдел (tubulus соntortus proximalis) в форме извитого канальца. Третий отдел сравнительно короток и прям. За ним вновь идет извитой четвертый отдел (tubulus со ntortus distalis), переходящий в пятый — прямую трубочку, впадающую в собирающие протоки мочеточника. Главным отличием от строения мочевых трубочек высших позвоночных является отсутствие петли Генле, которой топографически, но не функционально, отвечает третий отдел мочевого канальца лягушки. Именно отсутствием петли Генле и объясняется гипотоничность мочи лягушки. Размеры почечного тельца разных видов таковы:
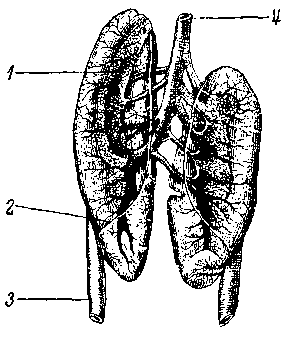
Рис. 1. Почки самца зеленой лягушки. Вид с брюшной стороны. Почечные артерии и выносящие сосуды семенников удалены:
1 — надпочечник, 2 — срединная граница брюшины, 3 — выносящий проток 4 — задняя полая вена.
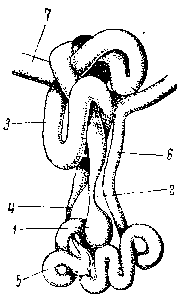
Рис. 2. Мочевая трубочка зеленой лягушки:
1— почечное тельце, 2 — первый отдел (шейка), 3 — второй отдел, 4 третий отдел, 5 — четвертый отдел, 6 — пятый отдел, 7 — собирательная трубочка мочеточника.
Виды | Длина (мм) | Ширина (мм) |
Лягушка-бык……. | 0, 125 | 0, 100 |
Зеленая ……….. | 0, 113 | 0, 088 |
Травяная ………. | 0, 074—0, 100 | 0, 055—0, 060 |
У лягушки-бык длина «шейки» 0, 100 мм, второго отдела 3, 80, третьего 0, 20, четвертого 2, 50 и пятого 0, 70.
Мочеточником служит выносящий проток (ductus deferens), соответствующий со сравнительно-анатомической точки зрения Вольфову протоку. Начальная часть каждого выносящего протока прилегает к наружному (боковому) краю соответствующей почки, причем самая передняя часть настолько погружена в вещество почки, что ее можно видеть только на поперечных срезах. Отделившись от заднего конца почки, выносящий проток направляется назад, прилегая к спинной стенке тела животного. Правый и левый протоки постепенно сближаются и заканчиваются рядом, но самостоятельно, на спинной стороне клоаки. У самцов задний участок каждого выносящего протока расширен в семенной пузырек (vesicula seminalis), особенно увеличенный во время размножения. В начале спаривания семенные пузырьки пусты, но потом наполняются спермой. Не исключена их внутрисекреторная функция. Связь со стремлением к спариванию теперь отрицается.
Мочевой пузырь (vesica urinaria), как уже указывалось выше, не связан непосредственно с мочевой системой. Это тонкостенный мешок, представляющий собой выпячивание брюшной стенки клоаки. Его основная часть несколько сужена и носит название «шейки», а свободная часть расширена и слегка двулопастна. В стенках мочевого пузыря имеется слой гладких мускульных волокон. Действие сжимателей клоаки может приводить отверстия выводных протоков почек к соприкосновению с отверстием мочевого пузыря. Этим путем мочевой пузырь наполняется мочей. Повидимому, главной его функцией является фильтрация воды, если организм начинает чувствовать в ней необходимость. Опорожнение мочевого пузыря происходит в результате совместного действия брюшных мускулов и мускульных волокон его собственных стенок. Контролируется этот процесс центром, находящимся в спинном мозгу на высоте пятого позвонка.
Каждый надпочечник (glandula suprarenalis) тесно связан как с брюшной поверхностью почки, так и с выносящими почечными венами. В свежем состоянии надпочечники хорошо отличаются своим золотисто-желтым цветом от красно-коричневых почек. Тело надпочечника плоско и лентообразно. Микроскопически можно различить в надпочечнике перемежающиеся эпителиальные более крупные и темные «мозговые», или хромафинные, клетки. В теплое время года добавляются еще особые «летние» клетки, а число «мозговых» клеток уменьшается. Для «летних» клеток особенно характерно сильное окрашивание рядом микроскопических красок. Функция надпочечника Лягушки совпадает с функцией надпочечника млекопитающих и сводится к выделению «мозговыми» клетками адреналина, поступающего в кровь. Последний, как известно, действует возбуждающе на периферические симпатические нервные окончания и дает сужение просвета кровяных сосудов. Одностороннее удаление надпочечника не дает у лягушек заметного эффекта (компенсация со стороны оставшегося), но двустороннее быстро приводит к смерти животного при явлениях отравления (параличи). Жизнь оперированной таким образом лягушки может быть поддержана пересадкой кусочков надпочечника в один из лимфатических мешков.
Половые органы самца
Органы размножения самца (organa genitalia virilia) состоят из парных семенников и их выводных протоков. Каждый семенник (testis) представляет собой овальное или округлое тело, несколько сжатое в спиннобрюшном направлении. Семенники зеленых лягушек чисто желтые и более округлые. У травяных лягушек они более продолговатые и более или менее пигментированы. Величина семенников меняется в течение года (наибольшая величина в августе, наименьшая в мае), но у травяной лягушки они всегда относительно крупнее, чем у зеленой. Помещаются семенники с брюшной стороны почек, будучи подвешены на особых брыжейках (mesorchium), и одеты, за исключением срединного края, серозным слоем брюшины.
На своей поверхности семенник одет белковой оболочкой (tunica albuginea), состоящей из плотной соединительной ткани с гладкими мускульными волокнами. Кроме сосудов и нервов, внутри семенника находится большое количество семенных трубочек (tubuli semi niferi), которые отделены друг от друга перегородками (septula testis), отходящими от белковой оболочки. Семенные трубочки выстланы семеобразователъным эпителием, изменяющимся в зависимости от времени года и могущим состоять из одного или более слоев.
Семенные трубочки постепенно собираются в семявыносящие канальцы (vasa efferentia testis), которые выходят в числе от 4 до 18 от срединного края семенника. Извиваясь в брыжейке семенника, эти канальцы достигают срединно-брюшного края почки и проникают в ее толщу. Сперматозоиды, или семенные нити, образовавшиеся в семенных трубочках, проходят через семенные канальцы в почку и затем попадают в выносящий проток почки, играющий, таким образом, у самцов роль семямочеточника (ductus uro-spermatiсus). Иногда у самцов позади или с наружной стороны почки наблюдается рудимент Мюллерова протока в виде тоненькой трубочки.
Внутри семенных трубочек сперматозоиды часто собраны пучками. У травяной лягушки процесс образования сперматозоидов носит более выраженный цикличный характер, нежели у зеленой. Поэтому в каждый данный момент в семенниках травяной лягушки можно найти сперматозоиды только одного возраста, тогда как у зеленой даже внутри одной семенной трубочки могут быть одновременно встречены разновозрастные сперматозоиды. Путем ряда последовательных делений первичные зародышевые клетки (сперматогонии) превращаются во вторичные (спгрматоциты), а видоизменением последних формируются сперматозоиды. Окончательно сформированные сперматозоиды состоят из головки и хвостика, имеющих различные размеры и форму у разных видов (рис. 220). Длина головки сперматозоидов зеленой лягушки колеблется от 0, 015 до 0, 021 мм при ширине от 0, 002 до 0, 003 мм.
Рис. 3. Мочеполовая система самца травяной лягушки во время размножения:
1— подпозвоночная оболочка, 2 — граница брюшины, 3 — семявыносящие канальцы, 4 — правая почка, 5 — граница брюшины, 6 — клоака, 7 — мускул сжиматель клоаки, 8 — артерии, 9 — жировое тело, 10 — левый семенник, 11 — выносящий проток, 12 — семенной пузырек, 13 — мочевой пузырь, 14 — прямая кишка.
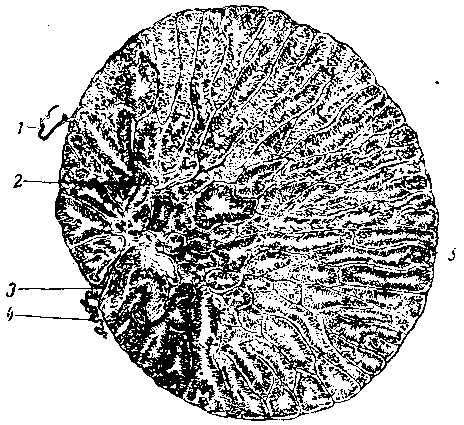
Рис. 4. Поперечный срез семенника травяной лягушки в начале размножения:
1 — брюшной листок брыжейки семенника, 2 – еыводной проток двух семенных трубочек, 3 — семявыносящий каналец, 4 — спинной листок брыжейки семенника, 5 — белковая оболочка.
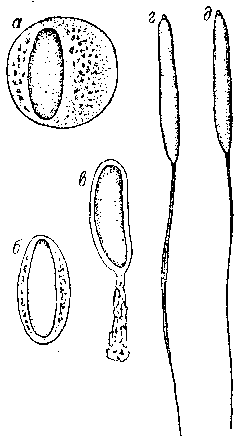
Рис. 5. Превращение сперматиды (а) зеленой лягушки в сперматозоид (д).
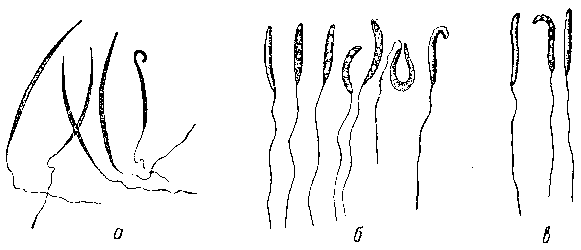
Рис. 6. Сперматозоиды травяной (а), остромордой (б) и зеленой (в) лягушек.
Длина хвостика в среднем около 0, 040 мм. Часто встречаются еще особые «гигантские» сперматозоиды с длиной головки в 0, 031 мм. У травяной лягушки сперматозоиды заметно крупнее и тоньше (длина головки 0, 051—0, 060, а ширина около 0, 001 мм).
Впереди семенников (у самки — яичников) располагаются желтые жировые тела (corpora adiposa). Каждое жировое тело, кроме мало заметной основной части, имеет еще от 3 до 12 лопастей. Жировые тела служат запасом материала для развития половых продуктов. В связи с этим они не изменяются почти во все время зимней спячки, но испытывают резкое уменьшение перед размножением. В течение лета наблюдается их увеличение. Сказанное понятно, если учесть, что голодание мешает половой диференциации неполовозрелых животных и вызывает вырождение половых клеток взрослых. Частичная кастрация дает увеличение жировых тел. Удаление последних вызывает вырождение половых продуктов, причем первыми погибают более законченные в своем развитии продукты.
Половые органы самки
Органы размножения самок (organa genitalia muliebrа) состоят из правого и левого яичников и двух яйцеводов.
Каждый яичник (ovarium) представляет собой тело, подвешенное на особой брыжейке (mesovariu m), с брюшной стороны по отношению к почкам. Снаружи яичник напоминает веер со многими складками, ибо его боковой край во много раз превосходит по длине срединный. Внутри яичника имеется щелевидная полость, выполненная особой жидкостью и поделенная поперечными перегородками на 6—7 изолированных карманов. Размеры яичника резко различаются в зависимости от времени года. После кладки он меньше всего, а осенью и особенно весной (у травяной лягушки с конца августа по апрель) занимает большую часть полости тела. В последнем состоянии просвечивающие яйца придают яичнику темную окраску. Стенки яичника состоят (снаружи внутрь) из слоя брюшного эпителия, среднего, или яйценосного, слоя (stratum medium=str. oophorum) и внутреннего эпителия. Обычно в стенках яичника наблюдаются яйца на разных стадиях развития (рис. 222). Яйцевые клетки (овоциты) окружены одним слоем фолликулярных клеток и соединительнотканной оболочкой (текой), содержащей гладкие мышечные волокна. Тека покрывает поверхность фолликула не полностью, оставляя непокрытой область будущего разрыва, непосредственно примыкающую к наружной стенке яичника. Молодые яйцевые клетки вначале прикрепляются к стенке яичника на широком пространстве, но, по мере роста, площадь их прикрепления постепенно сокращается. Рост яйцевых клеток сопровождается накоплением значительного количества желтка. Выделение яйцевой клетки (овуляция) слагается из разрыва фолликула и активного сокращения гладких мышечных волокон его соединительно-тканной оболочки. Последнее выталкивает яйцевую клетку через образовавшееся отверстие. Из полости тела яйцо через яйцеводы выводится наружу. Место разрыва спадается, и через некоторое время в стенке яичника остается лишь небольшое соединительнотканное утолщение. После овуляции всех крупных яйцеклеток старшего поколения яичники содержат только мелкие овоциты и окрашены в желтовато-коричневый цвет.
Если осенью или зимой ввести в спинной лимфатический мешок самки или в полость ее тела вещество нескольких лягушачьих нижних мозговых придатков (гипофизов), то через 20—48 часов наступает овуляция весеннего типа. Таким образом в лаборатории могут быть получены даже головастики (Кащенко, 1936, 1947).
В летние месяцы гипофиз травяной лягушки гонадотропного гормона не содержит.
Яйцевод (оvi ductus) лягушки со сравнительно-анатомической точки зрения представляет собой Мюллеров проток. Правый и левый яйцеводы построены одинаково и расположены довольно симметрично.
Каждый яйцевод представляет собой трубку, которая раз в 8 превышает длину тела взрослой самки. В нем можно различить три части: прямой (pars rесtа), извитой (pars convoluta) и задний отдел, или «матку» (pars utеri = «utеrus»). Первые два отдела иногда объединяются под именем «трубы» (tuba). Прямой отдел открывается в переднюю часть полости тела у основания легких брюшными устьями или «воронками» (ostium abdоminаlе), выстланными мерцательным эпителием. Извитой отдел яйцевода отличается железистым характером своих стенок. Один яйцевод лягушки может, разбухнув в воде, дать в среднем литр студенистой массы. Перед кладкой диаметр его заметно возрастает. Извитой отдел яйцевода прикрепляется к стенке полости тела особой брыжейкой (mesotubarium). Маточный отдел яйцевода более широк и имеет тонкие, растяжимые стенки. Каждый из яйцеводов (правый и левый) открывается в клоаку самостоятельно клоакальным отверстием (ostium сlоасаlе), расположенным на особом маточном сосочке (papilla uterina). Маточные сосочки лежат в клоаке до отверстий выводных каналов почек.
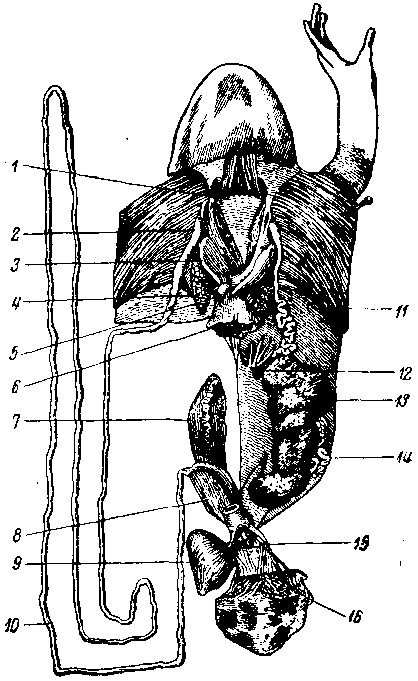
Рис. 7. Половые органы самки зеленой лягушки после икрометания (яичник мал, а яйцеводы узкие). Кишечник почти весь удален. Клоака надрезана и развернута. Правый яичник удален, а яйцевод расправлен:
1 — околосердечная сумка, 2 — прямой отдел яйцевода, 3 — печень, 4 — правое легкое, 5 — поджелудочная железа, 6—пищевод (перерезан), 7 — правая почка, 8 — правая матка, 9 — мочевой пузырь, 10 — извитой отдел яйцевода, 11— поперечный мускул, 12 — жировое тело, 13 — левый яичник, 14 — извитой отдел яйцевода, 15 — маточный сосочек, 16 — граница кожи и клоаки.
Яйца втягиваются брюшными отверстиями яйцеводов и продвигаются далее благодаря ресничкам мерцательного эпителия. В среднем отделе каждое яйцо обволакивается выделениями стенок, благодаря чему превращается нa выходе из тела лягушки в «икринку», ибо слизистые оболочки разбухают в воде. Готовое яйцо травяной лягушки, выпавшее в полость тела, весит в среднем 0, 002 г. Яйцо, взятое из яйцевода и одетое его еще не набухшими выделениями, весит в среднем 0, 0047 г. Маточный отдел яйцевода накапливает икринки и выпускает их наружу отдельными порциями.
Если у самцов жировые тела прилегают на значительном протяжении своей основной части к переднему краю семенника, то у самок жировые тела только заключены в брыжейку яичника, не касаясь его самого.
Гермафродитизм
Неоднократно у лягушек разных видов описывались различные аномалии половых органов и чаще всего гермафродитизм. Так, у зеленых лягушек были описаны гермафродитные особи (Фридман, 1898; Лавров, 1904, 1908), а Мелихова (1938) описала случай отсутствия половых желез. У чернопятнистой лягушки нашел гермафродитизм Тинг (1937). Особенно много наблюдений известно над гермафродитизмом травяных лягушек: Маршаль (1884), Педашенко (1890), Смит (1890), Латтер (1890), Коль (1896), Шмит-Марсель (1908), Воронцова (1926), Спауль и Керр (1940) и другие. Наконец, известны и случаи гермафродитизма лягушки-быка и Rana virescens.
Внешние факторы могут менять пол лягушек. Разумеется, воздействие на более ранних стадиях онтогенеза эффективнее, чем на последующих. У травяной лягушки в течение первого года после метаморфоза половые органы варьируют очень сильно. У осенних сеголеток определение пола даже при пом