Акт мочеиспускания рефлекторная дуга

^Нервная регуляция опорожнения мочевого пузыряМоча, постоянно вырабатывающаяся в почках, накапливается в мочевом пузыре, который периодически полностью опорожняется. Эта функция, играющая огромную роль в социальной жизни человека, опосредована как деятельностью гладких мышц мочевого пузыря, так и влияниями вегетативных и соматических нервов. Нервная регуляция функции мочевого пузыря заключается в чередова- ^ ЧАСТЬ IV. ПРОЦЕССЫ НЕРВНОЙ И ГУМОРАЛЬНОЙ РЕГУЛЯЦИИ
нии длительных периодов наполнения и коротких периодов опорожнения. Благодаря нервной активности опорожнение мочевого пузыря во время периода наполнения невозможно или затруднено. Скорость наполнения мочевого пузыря составляет примерно 50 мл в час. Вследствие пластичности гладких мышц пузыря (см. с. 86) давление в нем лишь незначительно повышается при увеличении объема (рис. 16.15). При накоплении приблизительно 150-250 мл мочи появляются первые короткие позывы к мочеиспусканию, обусловленные кратковременным повышением давления внутри пузыря. Период опорожнения обычно начинается, когда в мочевом пузыре накапливается порядка 250-500 мл мочи. Способность пузыря накапливать мочу называется удержанием мочи, а его опорожнение-актом мочеиспускания (рис. 16.15). Строение и иннервация мочевого пузыря (рис. 16.16). Мочевой пузырь представляет собой полый орган, образованный мышечными слоями; эти слои в совокупности называются изгоняющей мышцей (детрузором). Стенки пузыря состоят из сети длинных гладкомышечных волокон. В области основания пузыря находится треугольный участок, образованный тонкими гладкомышечными волокнами (пузырный треугольник). В углах основания этого треугольника расположены устья мочеточников. Мочеточники открываются в пузырь в косом направлении, поэтому при повышении внутрипузырного давления обратного заброса мочи в них не происходит. В области вершины треугольника отходит мочеиспускательный канал. Благодаря особому расположению мышц здесь образуется как бы функциональный сфинктер (внутренний сфинктер мочевого пузыря). При акте мочеиспускания внутренний сфинктер может расслабиться только в результате сокращения детрузора; вследствие сокращения этой мышцы мочеиспускательный канал под действием вплетающихся в него радиальных волокон укорачивается, что автоматически приводит к пассивному раскрытию внутреннего сфинктера. Мочеиспускательный канал замыкается, кроме того, наружным сфинктером, образованным поперечнополосатой мускулатурой промежности; у женщин этот сфинктер выражен слабо. Иннервация мочевого пузыря и сфинктеров схематически представлена на рис. 16.16. ^ тазовых нервов, отходящие от второго и четвертого крестцовых сегментов, возбуждают мускулатуру пузыря. Влияние этих нервов необходимо для нормальной регуляции акта мочеиспускания. Под действием симпатических нервов, отходящих от верхних поясничных сегментов, мышца, изгоняющая мочу, расслабляется, а мышцы пузырного треугольника сокращаются. Возможно, эти симпатические влияния способствуют удержанию мочи. Соматическая иннервация наружного сфинктера осуществляется двигательными волокнами срамного нерва, тела нейронов которых лежат в средних крестцовых сегментах. Информация о степени наполнения мочевого пузыря воспринимается рецепторами растяжения, расположенными в его стенке, и передается в центральную нервную систему по чувствительным волокнам тазовых нервов. Возможно, болевые и неболевые раздражения, возникающие при различных процессах в мочевом пузыре и мочеиспускательном канале, передаются в виде импульсов как в крестцовых, так и в поясничных висцеральных афферентах.
^
^ Из почечных лоханок моча поступает в мочевой пузырь благодаря перистальтическим сокращениям мочеточников. Чем более растянута стенка мочевого пузыря, тем сильнее возбуждаются расположенные в ней рецепторы растяжения. Возбуждение этих рецепторов приводит к активации парасимпатических нейронов, иннервирующих детрузор (рис. 16.17, рефлекторная дуга 1). В результате мочевой пузырь опорожняется. Как указано на рис. 16.17, дуга этого рефлекса замыкается на уровне передней области моста. Электрическое раздражение этой области приводит к опорожнению мочевого пузыря. С началом опорожнения пузыря процесс ускоряется лавинообразно до полного изгнания мочи. Этот протекающий по принципу положительной обратной связи (т.е. усиливающий сам себя) процесс, возможно, обусловлен следующими основными рефлексами: 1) сокращение мышцы, изгоняющей мочу, усиливает возбуждение афферентных волокон от рецепторов пузыря; 2) под влиянием струи мочи возбуждаются чувствительные волокна от рецепторов мочеиспускательного канала, что приводит к рефлекторной активации парасимпатических нервов мочевого пузыря; 3) происходит рефлекторное угнетение тормозных процессов в спинном мозге и вышележащих отделах. Все это сопровождается также рефлекторным расслаблением наружного сфинктера в результате торможения мотонейронов крестцовых сегментов. Как у животных, так и у людей перерыв спинного мозга выше крестцовых сегментов вначале приводит к полному подавлению рефлекторного опорожнения мочевого пузыря. Лишь по достижении хронической стадии заболевания (1-5 недель после травмы) вновь начинает действовать рефлекс опорожнения (автоматическое опорожнение). В этом случае рефлекторная дуга замыкается только в пределах спинного мозга (рис. 16.17, рефлекторная дуга 2). Вполне вероятно, что эта же дуга регулирует мочеиспускание у грудных детей; с возрастом же, по-видимому, этот рефлекс подавляется в результате тормозных влияний как от афферентных пузырных волокон, замыкающихся на уровне спинного мозга, так и от супраспинальных отделов. В течение первых дней или недель после возникновения параплегии или тетраплегии наблюдается вялость и атоничность мочевого пузыря. Если благодаря правильному уходу за больным не добавится инфекция мочевых путей, то начинается постепенный переход к автоматическому опорожнению мочевого пузыря. В этот период незначительное наполнение пузыря приводит к рефлекторному сокращению детрузора; мочеиспускание при этом учащено. В результате соответствующей тренировки такие больные могут управлять опорожнением пузыря. Они обучаются вызывать рефлекторные сокращения мышцы, изгоняющей мочу; для этого больной в определенное время, приуроченное к автоматическому ритму мочеиспускания, постукивает по нижнему отделу живота и надавливает на брюшную стенку, способствуя тем самым опорожнению пузыря (сегментарный рефлекс; см. с. 357). ^ Регуляция мочеиспускания и удержания мочи представляет собой в значительной степени автоматический рефлекторный процесс, однако на него оказывают влияние центры высших отделов ствола мозга, гипоталамус и большие полушария. Все эти влияния преимущественно ингибиторные, хотя некоторые из них могут быть и возбуждающими. О восходящих и нисходящих спинальных путях, соединяющих мочевой пузырь и мочеиспускательный канал с высшими отделами ЦНС, а также о расположении соответствующих нейронов в стволе мозга, гипоталамусе и коре больших полушарий известно мало. Деятельность высших отделов заключается, во-первых, в удержании мочи даже при сильном наполнении мочевого пузыря (это необходимо для того, чтобы мочеиспускание происходило только в надлежащих условиях) и, во-вторых, в произвольном мочеиспускании при наличии соответствующих позывов и условий [7, 11, 38]. Нарушения мочеиспускания весьма разнообразны и широко распространены. Непроизвольная задержка мочи может наступать при параличе или повреждении детрузора (например, при воспалительных заболеваниях или травматических повреждениях нервов), смешении мочеиспускательного канала (например, из-за опухоли предстательной 364 ^ железы) или при спазмах сфинктеров. Неспособность удерживать мочу называется недержанием. Она особенно часто встречается у рожавших женщин (например, в результате опущения матки из-за слабости мышц дна таза), а также при органических поражениях мозга (например, при рассеянном склерозе или атеросклерозе сосудов мозга у пожилых людей). Встречается также психогенное недержание мочи [16]. ^ Опорожнение кишечника (дефекация) и удержание каловых масс представляют собой важнейшие функции прямой кишки и ануса. Эти функции регулируются энтеральной нервной системой, парасимпатическими нервами из крестцовых сегментов и соматомоторными нервными влияниями. Внутренний анальный сфинктер имеет плотную симпатическую иннервацию, однако о роли симпатических нервов в функции низших отделов кишечника известно мало. Удержание кала. В области дистального конца прямой кишки имеются два сфинктера. ^ образованный гладкими мышцами, управляется непроизвольно. Наружный анальный сфинктер состоит из поперечнополосатых мышечных волокон, иннервируемых мотонейронами крестцового отдела (S2-S4), отростки которых идут в составе тазовых нервов. В покое оба этих сфинктера замкнуты. Тоническое сокращение наружного сфинктера поддерживается в результате спинальных рефлексов, афферентное звено которых включает импульсы от мышечных волокон сфинктера и окружающих тканей, и особенно кожи области анального отверстия. Когда в результате перистальтических сокращений нисходящей толстой кишки каловые массы поступают в прямую кишку, стенки ее растягиваются, что приводит к расслаблению внутреннего анального сфинктера и усилению сокращения наружного сфинктера. Расслабление внутреннего сфинктера связано главным образом с рефлексом, замыкающимся в пределах энтеральной нервной системы. Сокращение наружного сфинктера также возникает рефлекторно под влиянием сигналов, поступающих в крестцовые отделы спинного мозга по афферентным волокнам тазовых нервов (рис. 16.18). Эти процессы сопровождаются появлением позывов к дефекации – осознанных ощущений, связанных с возбуждением рецепторов стенок толстой и прямой кишок. Через несколько десятков секунд внутренний сфинктер вновь постепенно сокращается и наступает адаптация стенок прямой кишки к ее увеличенному объему. Эта адаптация обусловлена пластичностью ректальной мускулатуры. Напряжение в стенке прямой кишки снижается, и позыв к дефекации стихает. Благодаря всем этим процессам, находящимся под
контролем нервных влияний, здоровый человек может сдерживать позывы к дефекации до тех пор, пока объем содержимого прямой кишки не превысит 2 л. Важнейшую роль в удержании каловых масс играют супраспинальные центры, и особенно кора головного мозга: эти отделы оказывают возбуждающее влияние на мотонейроны наружного сфинктера и, возможно, ингибирующее влияние на парасимпатические спинальные рефлексы. Дефекация. Опорожнение прямой кишки в норме совершается произвольно. Под действием супраспинальных облегчающих влияний на спинальные парасимпатические рефлексы нисходящая, сигмовидная и прямая кишка (особенно их продольная мускулатура) сокращаются. Одновременно наступает расслабление обоих сфинктеров. Важным условием дефекации является повышение внутрибрюшного давления под действием, во-первых, усиленных сокращений мышц брюшной стенки и, во-вторых, опущения диафрагмы, наступающего в результате сокращения мышц грудной клетки, находящейся в положении вдоха, при замкнутой голосовой щели. Все эти механизмы приводят к тому, что тазовое дно опускается и каловые массы, содержащиеся в нисходящей, сигмовидной и прямой кишке, выбрасываются. Рефлексы дефекации полностью исчезают при разрушении крестцовых сегментов спинного мозга. После перерыва спинного мозга выше этих сегментов спинальные рефлексы дефекации сохраняются, однако произвольные движения, способствующие изгнанию каловых масс, не могут осуществляться. ^ Правда, они могут быть компенсированы (например, расширением наружного анального сфинктера руками), и благодаря этому больные параплегией способны регулярно опорожнять кишечник [7, 50]. | ||||||||
| Физиология человека. В 3-х томах. Т. Пер с англ./ под ред. Р. Шмидта и Г. Тевса. М.: Мир, 1996. 198 с Физиология человека: в 3-х томах / пер с англ под ред. Р. Шмидта и Г. Тевса. – М.: Мир. 1996.] | Руководство по терапевтической гомеопатии: Пер с англ. М.: Атлас, 1994. 205 с Аллен Х. К. Основания и показания к назначению и характеристики ведущих гомеопатичекских препаратов и нозодов со сравнением их патогенезов:… | |||||||
| Література … | Глоссарий Глоссарий взят из книги Брамм П., Брамм Д. Микропроцессор 80386 и его программирование: Пер с англ. М.: Мир, 1990,-448 с. Абсолютный адрес Глоссарий взят из книги Брамм П., Брамм Д. Микропроцессор 80386 и его программирование: Пер с англ. М.: Мир, 1990,-448 с | |||||||
| Пустыльник Е. И. Статистические методы анализа обработки наблюдений Базара М., Шетти К. Нелинейное программирование. Теория и алгоритмы: Пер с англ. М.: Мир, 1982. – 583 С | Список рекомендованої літератури Дункан Д. У. Основополагающие идеи в менеджменте. Уроки основоположников менеджмента и управленческой практики / Пер с англ. – М.:… | |||||||
| Список рекомендованої літератури Дункан Д. У. Основополагающие идеи в менеджменте. Уроки основоположников менеджмента и управленческой практики / Пер с англ. – М.:… | О.І. Рогач [та ін.]; pа ред. О.І. Рогача. К.: Либідь,2003. 784с. Isbn 966-06-0293-6 Мировые финансы[Текст]: пер с англ./ М. В. Энг, Ф. А. Лис, Л. Д. Мауер. М.: ДеКА,1998. 736с. Алф указ.: с. 722-734. Isbn 5-89645-004-4(рус.):… | |||||||
| Учебник для вузов системы мвд/ : в 2-х кн под ред проф. А. А. Пушкина (Олександр Анатолійович). Х.: Основа,1996 Цивільне право України : Підручник : у 2-х кн. /за ред. О. В. Дзери, Н. С. Кузнєцової-К.: Юрінком Інтер, 2001; 2002, 2004 | Руководство по коксованию том пер с немецкого Продукты каменноугольной смолы из книги: “Руководство по коксованию” том пер с немецкого под ред. О. Гросскинского с. 437 М.: Металлургия,… |
Источник
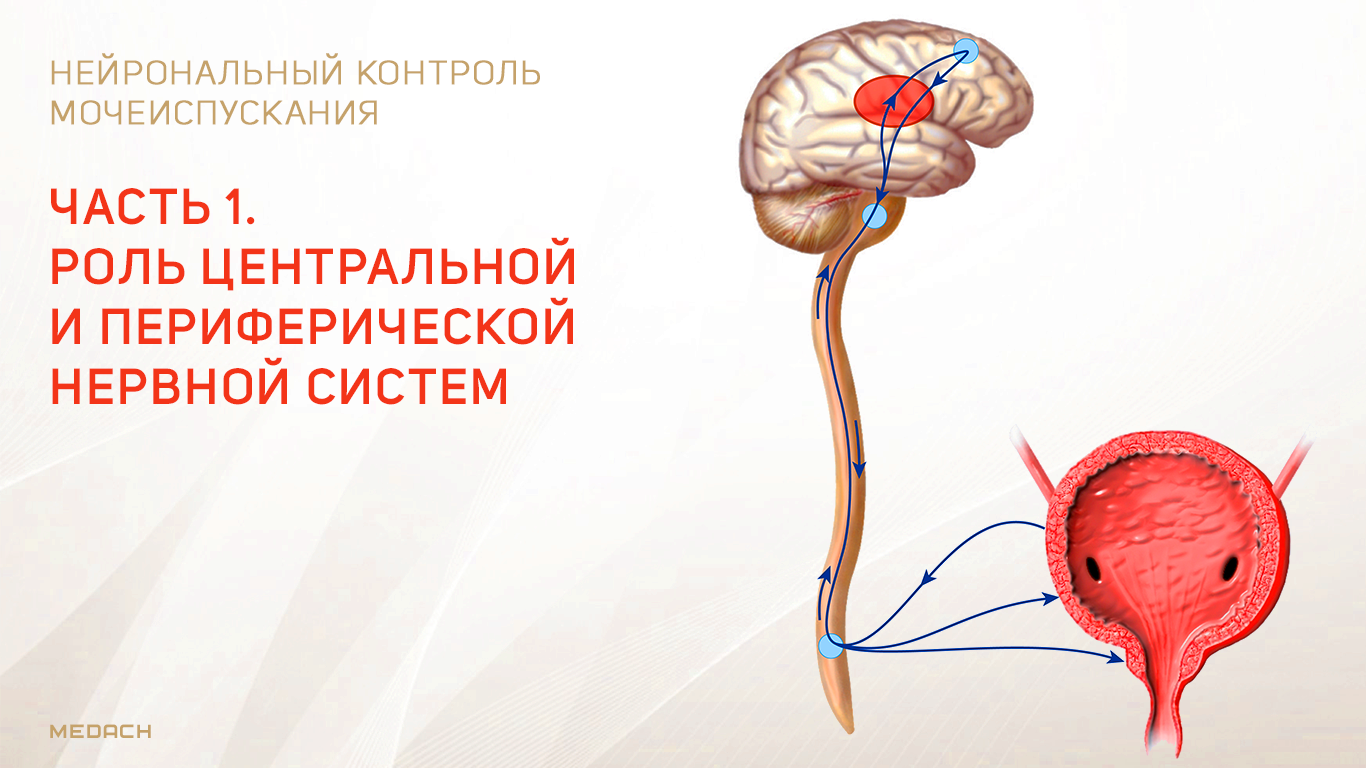
Хранение мочи и периодическое опорожнение мочевого пузыря (МП) зависят от координированной работы гладких и поперечнополосатых мышц, условно разделенных на два структурно функциональных отдела: резервуара (мочевой пузырь) и выпускной системы, состоящей из шейки МП, уретры и наружного сфинктера мочеиспускательного канала. За координированную работу вышеперечисленных структур отвечают сложные нейронные системы управления, расположенные в периферических ганглиях, спинном и головном мозге.
Обеспечение сознательного контроля мочеиспускания требует сложных взаимодействий между вегетативным (симпатика и парасимпатика) и соматическим отделами нервной системы.
Симпатические нервные волокна берут свое начало из боковых рогов грудопоясничного отдела (спинномозговой центр Якобсона) и ганглиях нижнего брыжеечного сплетения и достигают МП в составе подчревного нерва. Симпатические постганглионарные нервы выделяют норадреналин, который активирует β-адренергические рецепторы, ингибирующие сокращение детрузорной мышцы, и α-адренергические рецепторы, возбуждающие мускулатуру уретры и шейки МП. Симпатическое воздействие приводит к расслаблению мышцы, изгоняющей мочу, сокращению мускулатуры уретры и шейки МП и ингибированию интрамуральных ганглиев МП.
Центральный отдел парасимпатической иннервации МП располагается в промежуточных ядрах крестцовых сегментов. Холинергические преганглионарные волокна из промежуточных ядер посылают свои аксоны через тазовые нервы к ганглиозным клеткам тазового сплетения и интрамуральным нейронам в стенке мочевого пузыря. Ганглиозные клетки в свою очередь возбуждают детрузор мочевого пузыря, что приводит к его сокращению с последующим опорожнением. Основными медиаторами парасимпатической нервной системы тут являются ацетилхолин и другие нехолинергические медиаторы. Ацетилхолин действует опосредованно, возбуждая M3-холинорецепторы клеток детрузора. Парасимпатические нервные окончания в нервно-мышечных синапсах и в парасимпатических ганглиях тоже имеют холинорецепторы, возбуждение этих рецепторов на нервных окончаниях может усиливать (через рецепторы М1) или подавлять (через рецепторы М4) высвобождение медиаторов в зависимости от интенсивности нейронного возбуждения. Основной нехолинергический медиатор это АТФ, который активирует внутриклеточную пуринергическую систему через возбуждение P2X рецепторов и тоже способствует сокращению детрузора. Парасимпатические волокна вызывают расслабление ГМК уретры путем высвобождения оксида азота (NO).
Аксоны соматических двигательных нейронов передних рогов крестцовых сегментов S2-S4 (ядро Онуфа) проходят в срамном нерве и иннервируют поперечно-полосатые мышцы наружного уретрального сфинктера. Нейроны более медиально расположенного моторного ядра на том же уровне позвоночника иннервируют мускулатуру тазового дна.
Афферентные пути НМП состоят из цепочек чувствительных нейронов. Первые нейроны, располагающиеся в спинальных ганглиях на уровне S2-S4 и T11-L2, реагируют на пассивное растяжение и активное сокращение мышц МП и передают эту информацию на нейроны второго и третьего порядка. Эти нейроны обеспечивают координированную работу спинальных рефлексов и восходят к вышележащим структурам головного мозга, контролирующих фазы накопления и опорожнения МП. Наиболее важные афферентные волокна от мочевого пузыря идут в составе тазового нерва, в то время как чувствительность от шейки МП и уретры передается по срамным и подчревным нервам. Афферентные волокна этих нервов состоят из миелинизированных (Аδ) и немиелинизированных (С) аксонов. Aδ-волокна передают информацию о наполнении мочевого пузыря. С-волокна нечувствительны к изменению объема мочевого пузыря в физиологических условиях, поэтому они называются «тихими». Они реагируют главным образом на патологические стимулы, такие как химическое раздражение или охлаждение.
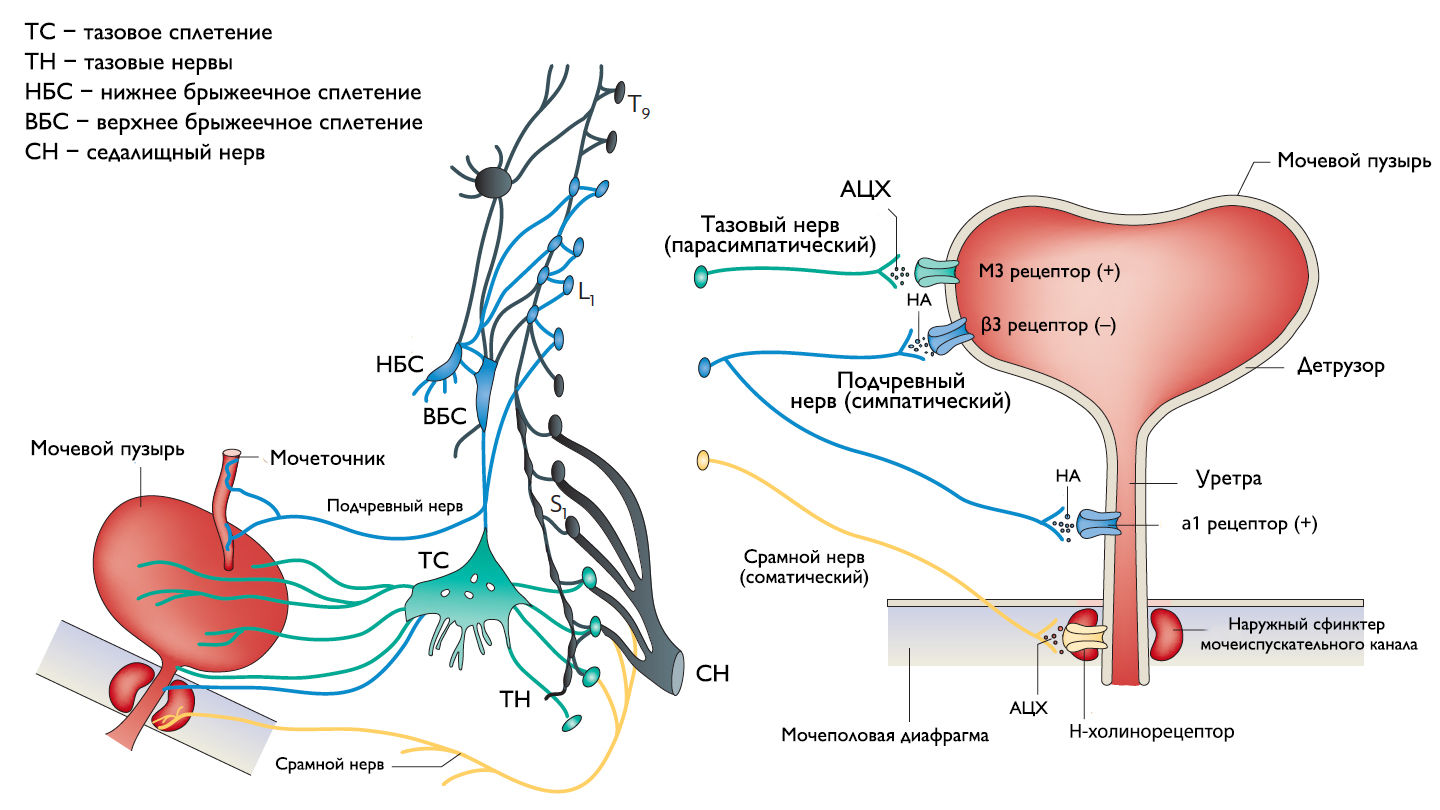
Рисунок 1 | Иннервация нижних мочевыводящих путей.
Специфический и неспецифический восходящий супраспинальный сенсорный путь
Одни спинномозговые промежуточные нейроны посылают восходящие волокна к определенным областям моста и среднего мозга, участвующим в мочеиспускании. Другие промежуточные нейроны передают информацию из нижних мочевыводящих путей в структуры переднего мозга, включая таламус и гипоталамус. Спиноталамический и спиногипоталамический тракты хотя и не играют главную роль в мочеиспускании, но могут включаться в сознательный контроль полноты мочевого пузыря. Чувствительные зоны коры ГМ через спиноталамический тракт информируются о состоянии наполнения МП.
Мостовой центр мочеиспускания (МЦМ) и его нисходящие спинномозговые двигательные пути
Впервые центр управления мочеиспусканием был открыт в дорсальной части моста Баррингтоном в 1925 году и с тех пор называется мостовым центром мочеиспускания или ядром Баррингтона. МЦМ располагается в области покрышки моста. Нейроны МЦМ имеют нисходящие возбуждающие синаптические контакты с клетками парасимпатических преганглионарных мотонейронов, иннервирующих постганглионарные клетки мочевого пузыря. Электрическая и химическая стимуляция МЦМ у крыс и кошек инициирует сокращение мочевого пузыря и расслабляет сфинктер уретры имитируя нормальное мочеиспускание. Цикл рефлекса мочеиспускания состоит из трех фаз, контролируемых различными центральными механизмами: фаза реализации безопасной среды – для начала мочеиспускания человеку необходимо осознание, что окружающие обстановка комфортна; фаза релаксации наружного уретрального сфинктера; и фаза сокращения мышцы, выталкивающей мочу. Процесс нормального мочеиспускания невозможен без какой-либо из этих фаз. МЦМ является командным центром мочеиспускания, который контролирует последовательное переключение фазы расслабления наружного уретрального сфинктера на фазу сокращения детрузорной мышцы.
Мостовой центр удержания мочи (МЦУ) и его нисходящие спинномозговые двигательные пути
Его роль заключается в расслаблении детрузора и сокращении наружного уретрального сфинктера. МЦУ располагается вентролатеральней МЦМ. Синапсы волокон МЦУ возбуждают ядро Онуфа в крестцовых сегментах спинного мозга, повышая таким образом тонус наружного сфинктера уретры. Стимуляция области МЦУ останавливает мочеиспускание, возбуждает мышцы тазового дна и сокращает уретральный сфинктер. Наоборот, двусторонние поражения МЦУ вызывают недержание мочи, чрезмерную детрузорную активность, невозможность хранения мочи и снижение тонуса уретрального сфинктера. На сегодняшний день нет анатомических доказательств связи между МЦУ и МЦМ, и было высказано предположение, что эти центры функционально независимы.
Роль кортикальных областей
Наиболее частые симптомы поражения кортикальных областей ГМ это поллакиурия и ургентное недержание мочи. Поэтому Andrew и Nathan выдвинули гипотезу, что отсоединение лобной или передней поясной извилины от гипоталамуса приводит к непроизвольному началу мочеиспускания [3]. Действительно, префронтальная кора головного мозга человека и передняя поясная извилина активируются во время мочеиспускания [4].
Мозжечок и базальные ганглии
Существует ряд исследований о том, что мозжечок и базальные ганглии оказывают в основном ингибирующее действие на мочевой пузырь. Мозжечковая патология приводит к увеличению частоты мочеиспускания и ургентному недержанию мочи. Симптомы гиперактивного мочевого пузыря также встречаются при болезни Паркинсона. Поскольку нет прямых связей этих областей с МЦМ, ингибирующее влияние, вероятно, косвенное через структуры переднего и среднего мозга.
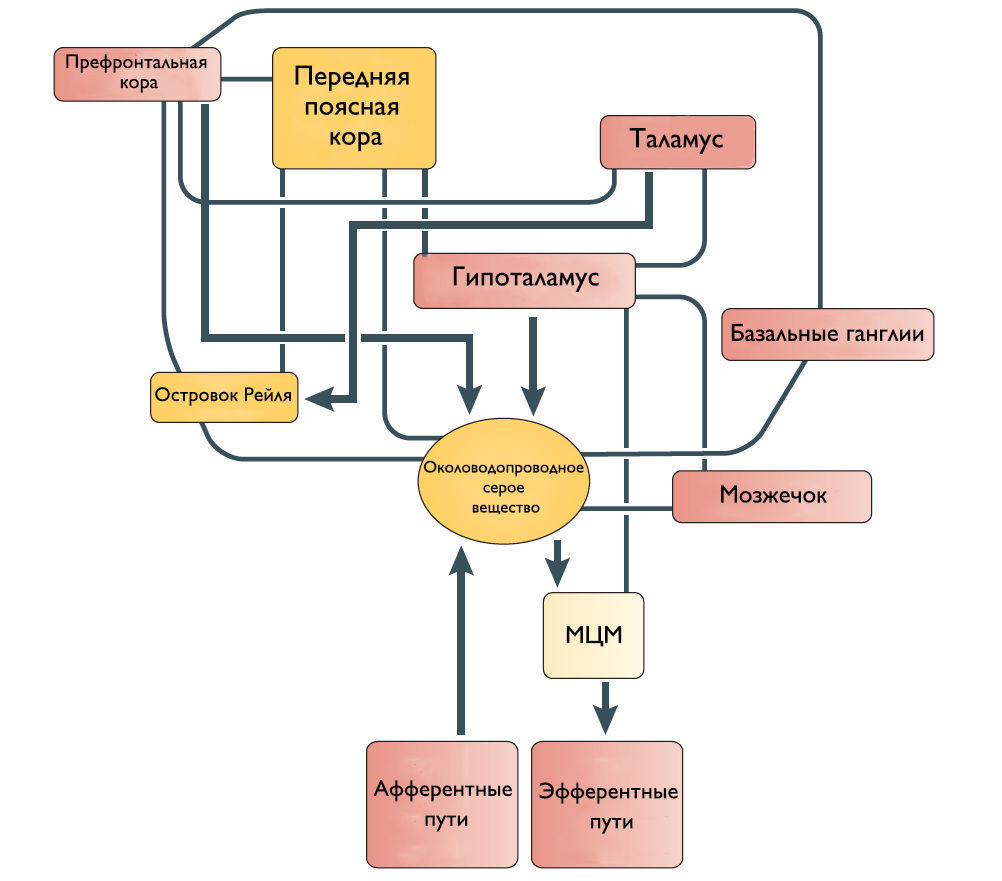
Рисунок 2 | Предположительное схематическое изображение связей между различными структурами переднего мозга и ствола мозга, которые участвуют в контроле мочеиспускания.
- Liao L., Madersbacher H. (ed.). Neurourology: Theory and Practice. – Springer, 2019.
- Clare J. Fowler et al. The neural control of micturition. – Nature Reviews | Neuroscience, volume 9. – June 2008.
- Andrew J, Nathan PW. Lesions of the anterior frontal lobes and disturbances of micturition and defecation. Brain. 1964;87:233-62.
- Griffiths, Derek J. “Use of al imaging to monitor central control of voiding in humans.” Urinary Tract. Springer, Berlin, Heidelberg, 2011. 81-97.
Нашли опечатку? Выделите фрагмент и нажмите Ctrl+Enter.
Источник